

Cell Type-Specific Transcriptomics Reveals that Mutant Huntingtin Leads to Mitochondrial RNA Release and Neuronal Innate Immune Activation
- PMID: 32681824
- PMCID: PMC7486278
- DOI: 10.1016/j.neuron.2020.06.021
Cell Type-Specific Transcriptomics Reveals that Mutant Huntingtin Leads to Mitochondrial RNA Release and Neuronal Innate Immune Activation
Abstract
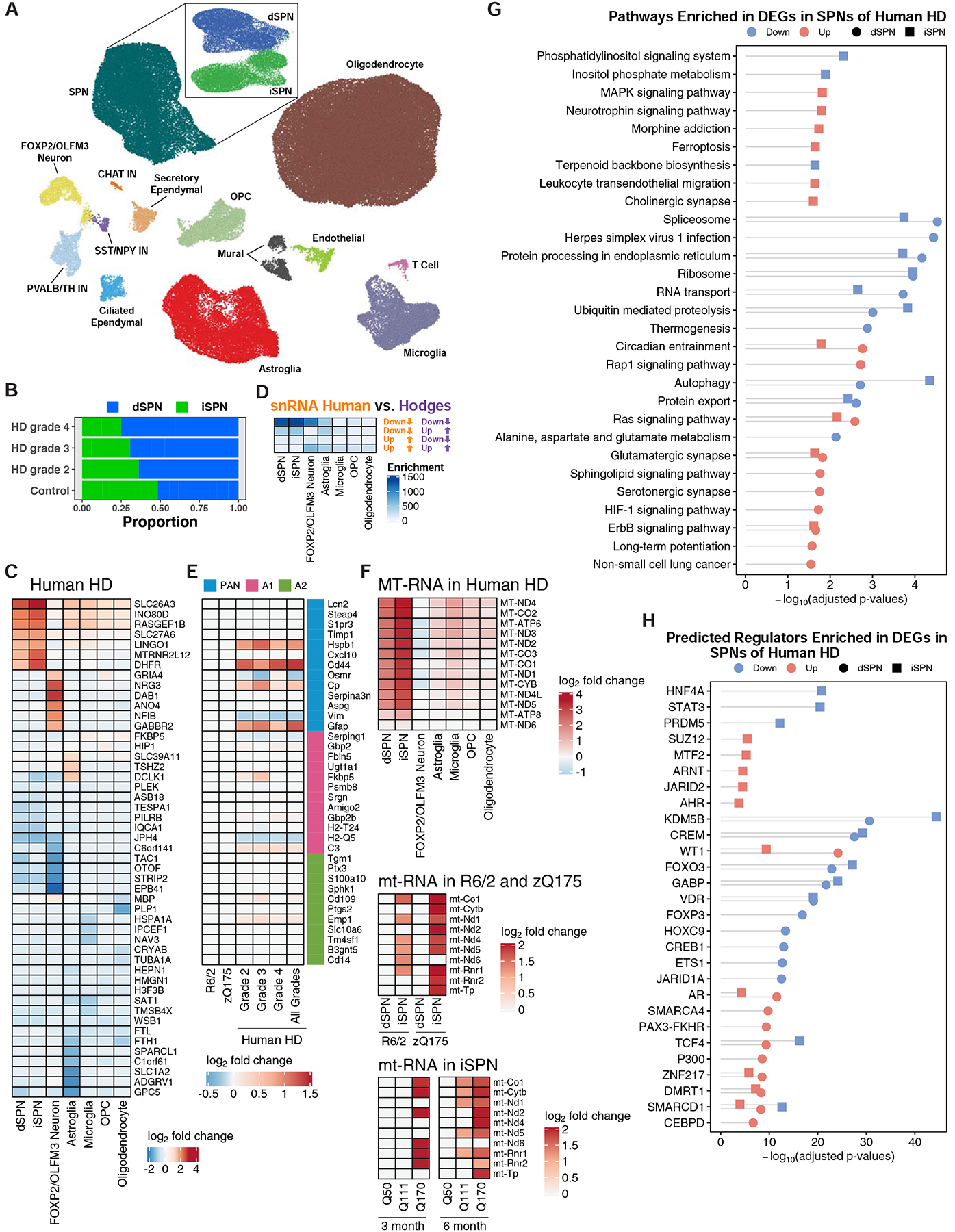
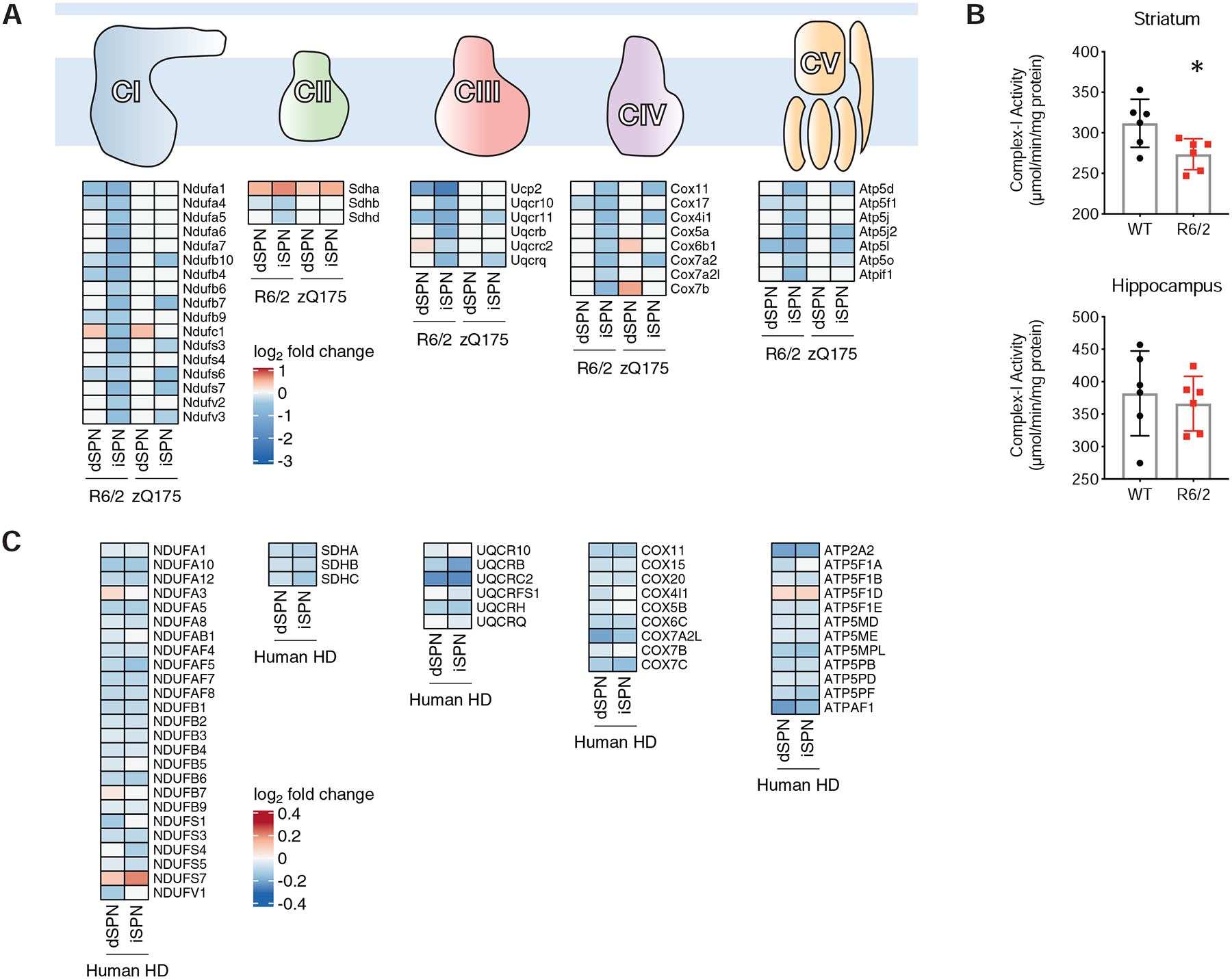
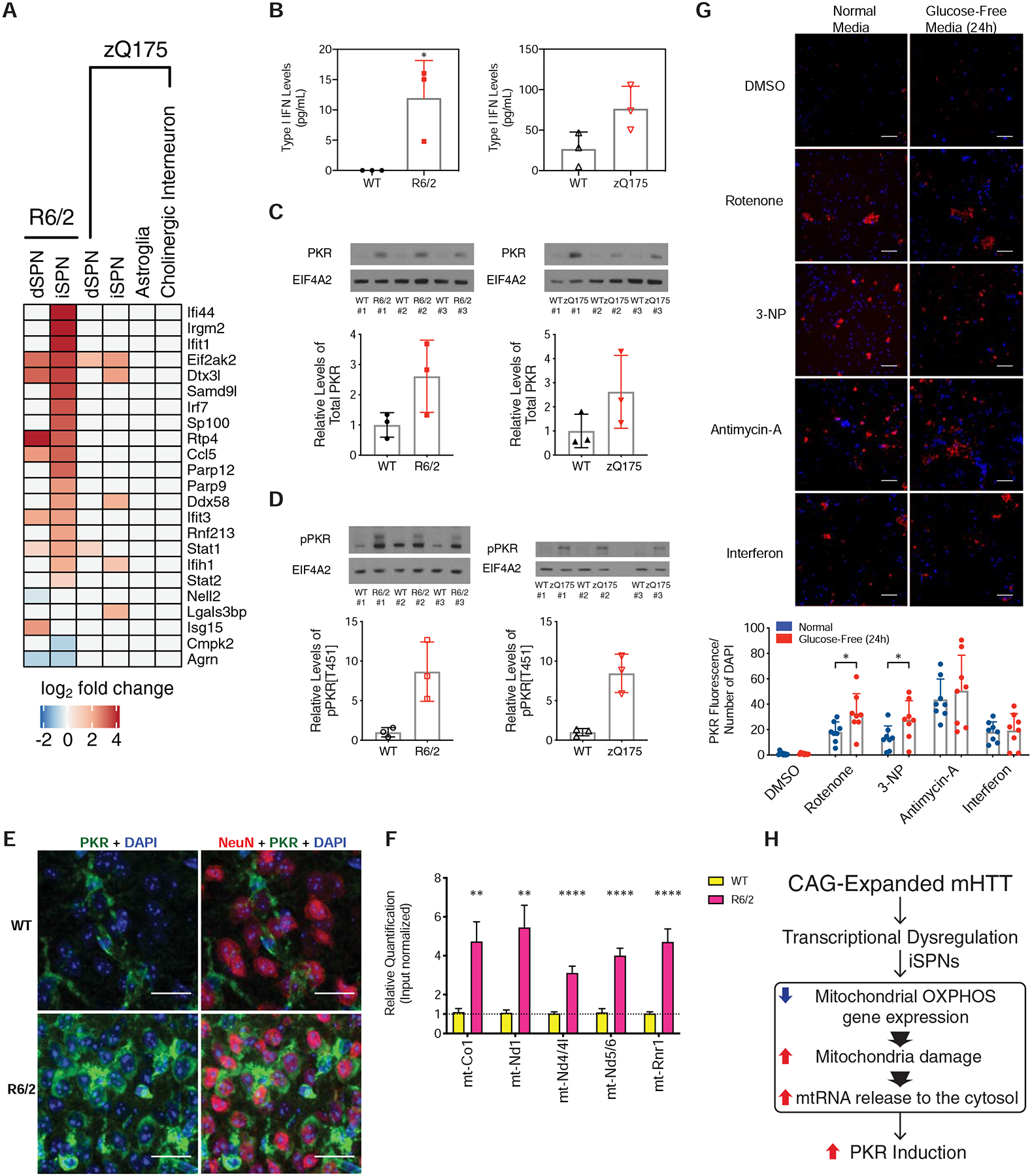
The mechanisms by which mutant huntingtin (mHTT) leads to neuronal cell death in Huntington's disease (HD) are not fully understood. To gain new molecular insights, we used single nuclear RNA sequencing (snRNA-seq) and translating ribosome affinity purification (TRAP) to conduct transcriptomic analyses of caudate/putamen (striatal) cell type-specific gene expression changes in human HD and mouse models of HD. In striatal spiny projection neurons, the most vulnerable cell type in HD, we observe a release of mitochondrial RNA (mtRNA) (a potent mitochondrial-derived innate immunogen) and a concomitant upregulation of innate immune signaling in spiny projection neurons. Further, we observe that the released mtRNAs can directly bind to the innate immune sensor protein kinase R (PKR). We highlight the importance of studying cell type-specific gene expression dysregulation in HD pathogenesis and reveal that the activation of innate immune signaling in the most vulnerable HD neurons provides a novel framework to understand the basis of mHTT toxicity and raises new therapeutic opportunities.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests M.H. is a member of the Hereditary Disease Foundation’s Scientific Advisory Board, and T.V. and H.W. are employed by CHDI Management, Inc., as advisors to the CHDI Foundation, Inc.
Figures
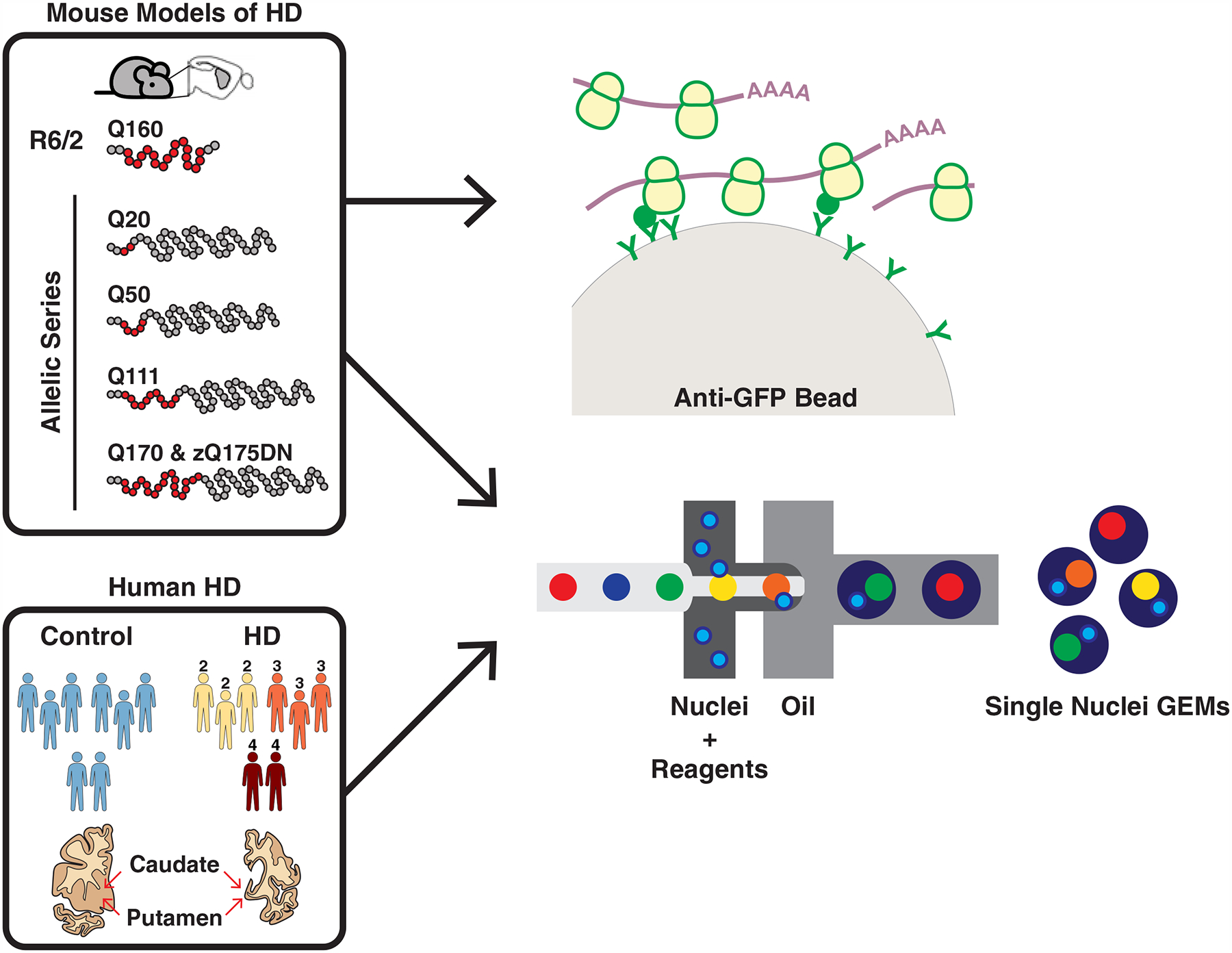
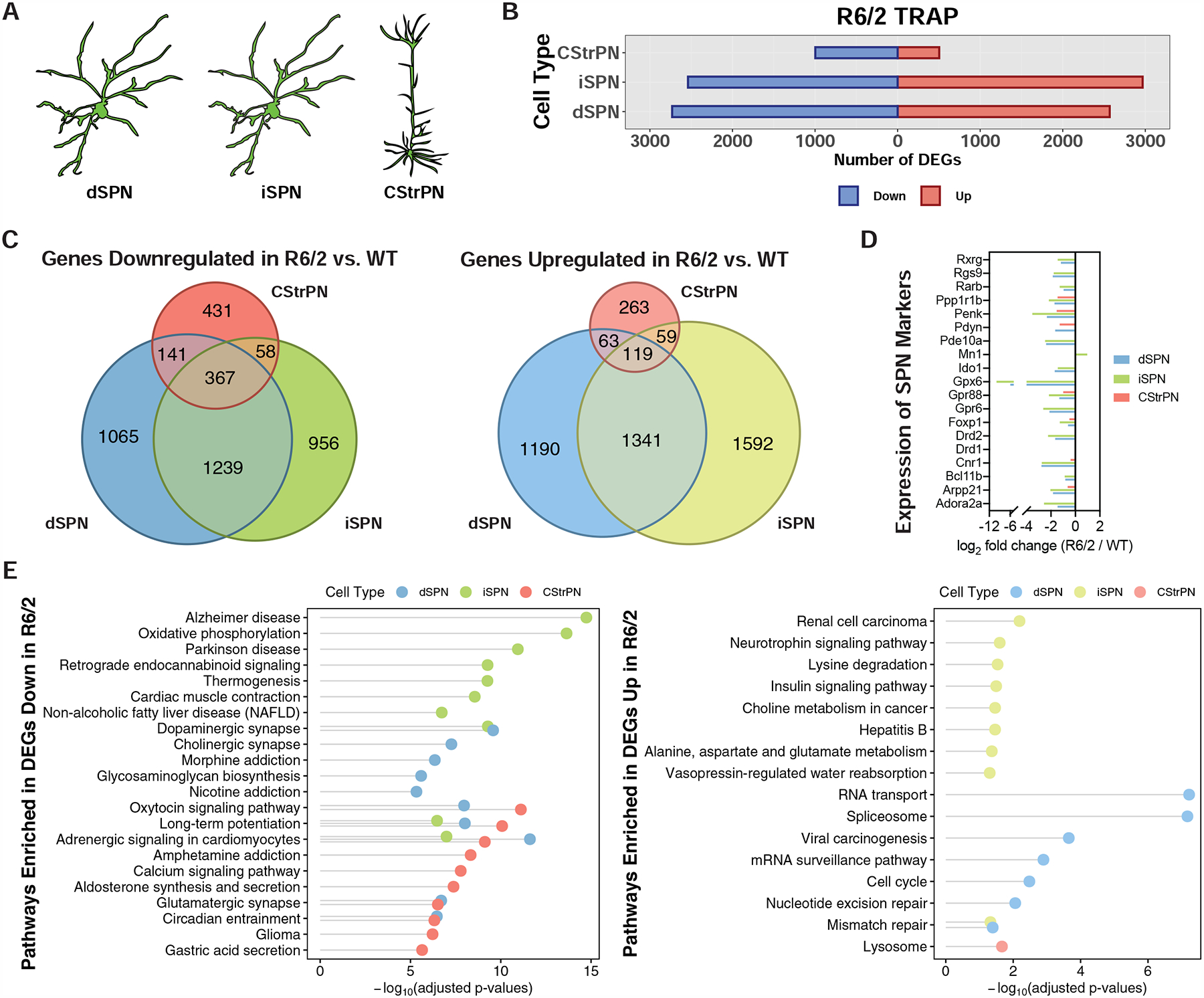
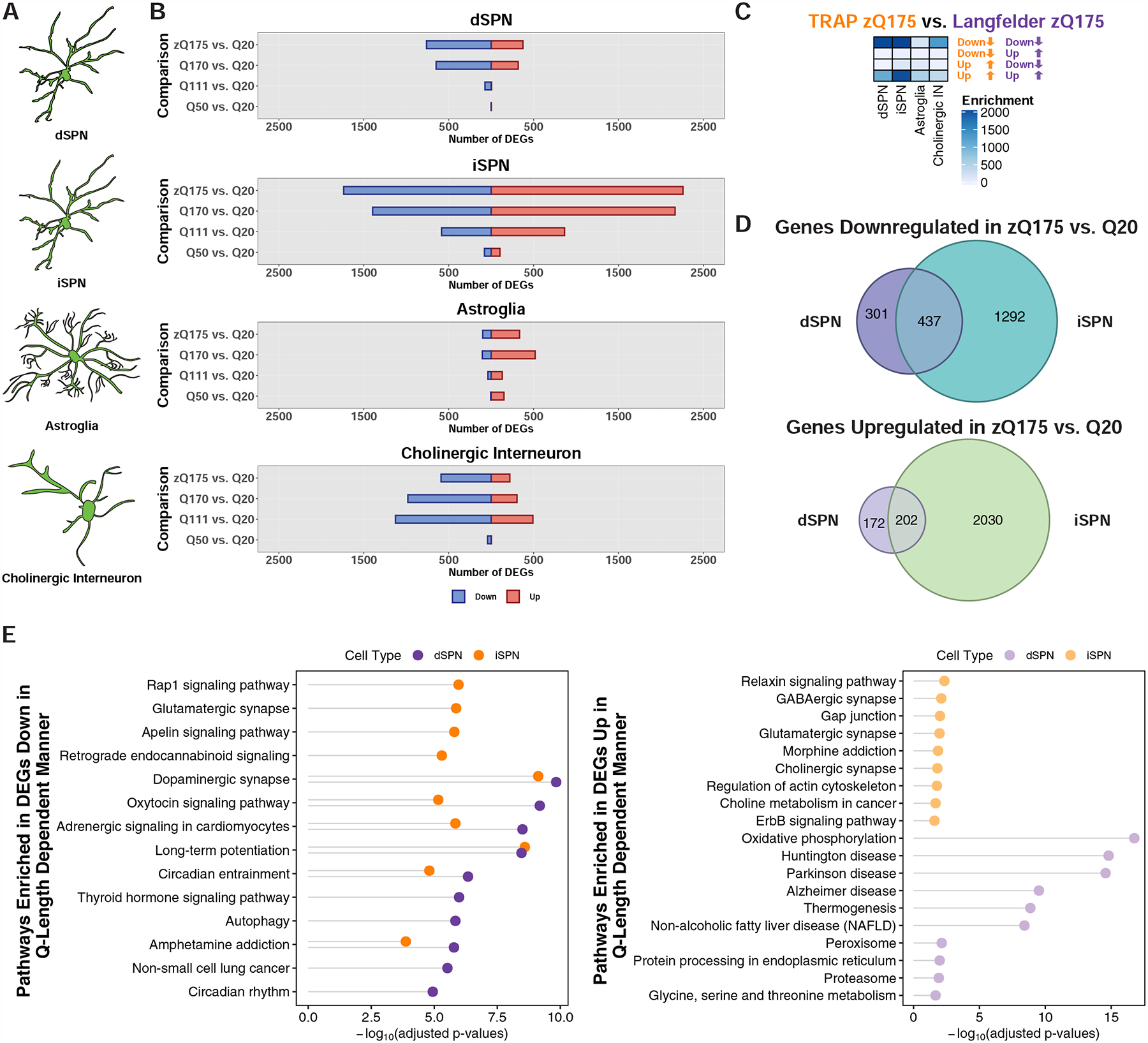
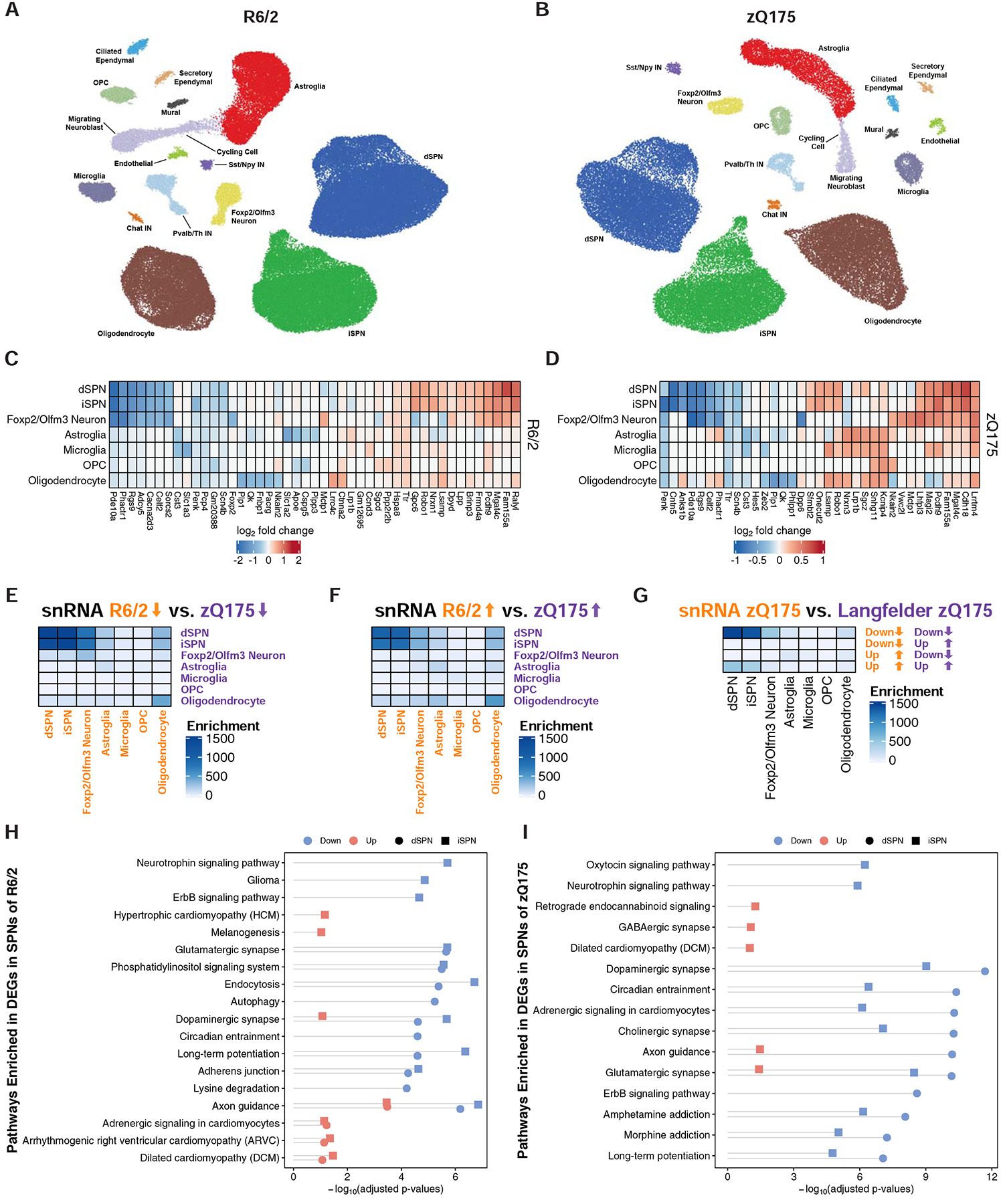
Comment in
-
Single Cell Technologies Define New Therapeutic Avenues for Huntington's Disease.Neuron. 2020 Sep 9;107(5):768-769. doi: 10.1016/j.neuron.2020.08.007. Neuron. 2020. PMID: 32910889
References
-
- Albin RL, Reiner A, Anderson KD, Dure L.S.t., Handelin B, Balfour R, Whetsell WO Jr., Penney JB, and Young AB (1992). Preferential loss of striato-external pallidal projection neurons in presymptomatic Huntington’s disease. Annals of neurology 31, 425–430. - PubMed
-
- Alfadhel M, Almuntashri M, Jadah RH, Bashiri FA, Al Rifai MT, Al Shalaan H, Al Balwi M, Al Rumayan A, Eyaid W, and Al-Twaijri W (2013). Biotin-responsive basal ganglia disease should be renamed biotin-thiamine-responsive basal ganglia disease: a retrospective review of the clinical, radiological and molecular findings of 18 new cases. Orphanet journal of rare diseases 8, 83. - PMC - PubMed
-
- Becht E, McInnes L, Healy J, Dutertre CA, Kwok IWH, Ng LG, Ginhoux F, and Newell EW (2018). Dimensionality reduction for visualizing single-cell data using UMAP. Nature biotechnology. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- RF1 AG054012/AG/NIA NIH HHS/United States
- R01 NS110188/NS/NINDS NIH HHS/United States
- R25 MH101076/MH/NIMH NIH HHS/United States
- T32 AG000222/AG/NIA NIH HHS/United States
- R01 NS065013/NS/NINDS NIH HHS/United States
- T32 EB019940/EB/NIBIB NIH HHS/United States
- U01 NS110453/NS/NINDS NIH HHS/United States
- R01 AG062335/AG/NIA NIH HHS/United States
- R01 MH109978/MH/NIMH NIH HHS/United States
- U01 MH119509/MH/NIMH NIH HHS/United States
- R01 AG058002/AG/NIA NIH HHS/United States
- R01 HG008155/HG/NHGRI NIH HHS/United States
- RF1 AG062377/AG/NIA NIH HHS/United States
- R01 NS100802/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
