

A Neural Network for Wind-Guided Compass Navigation
- PMID: 32681825
- PMCID: PMC7507644
- DOI: 10.1016/j.neuron.2020.06.022
A Neural Network for Wind-Guided Compass Navigation
Abstract
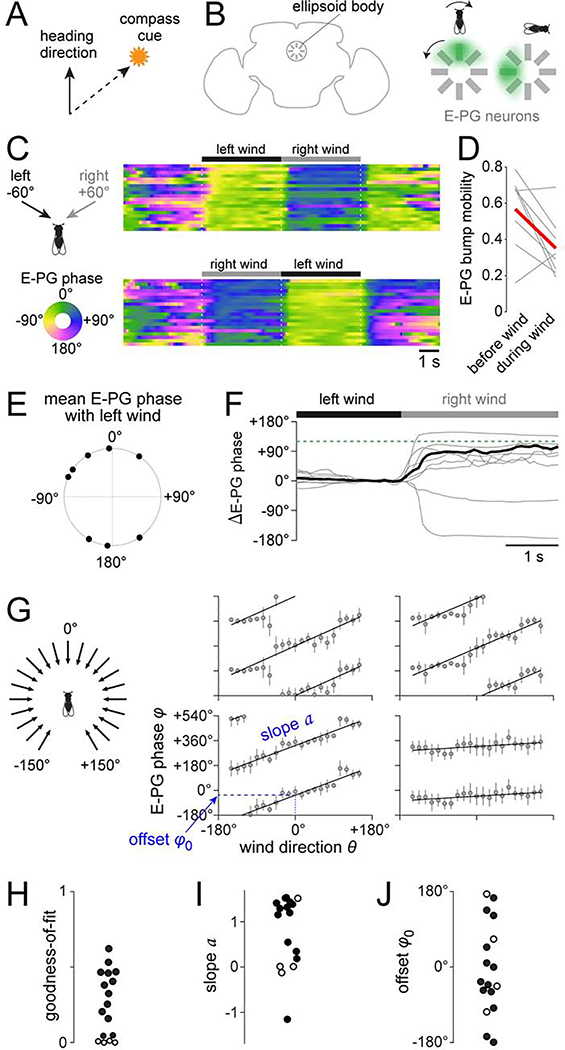
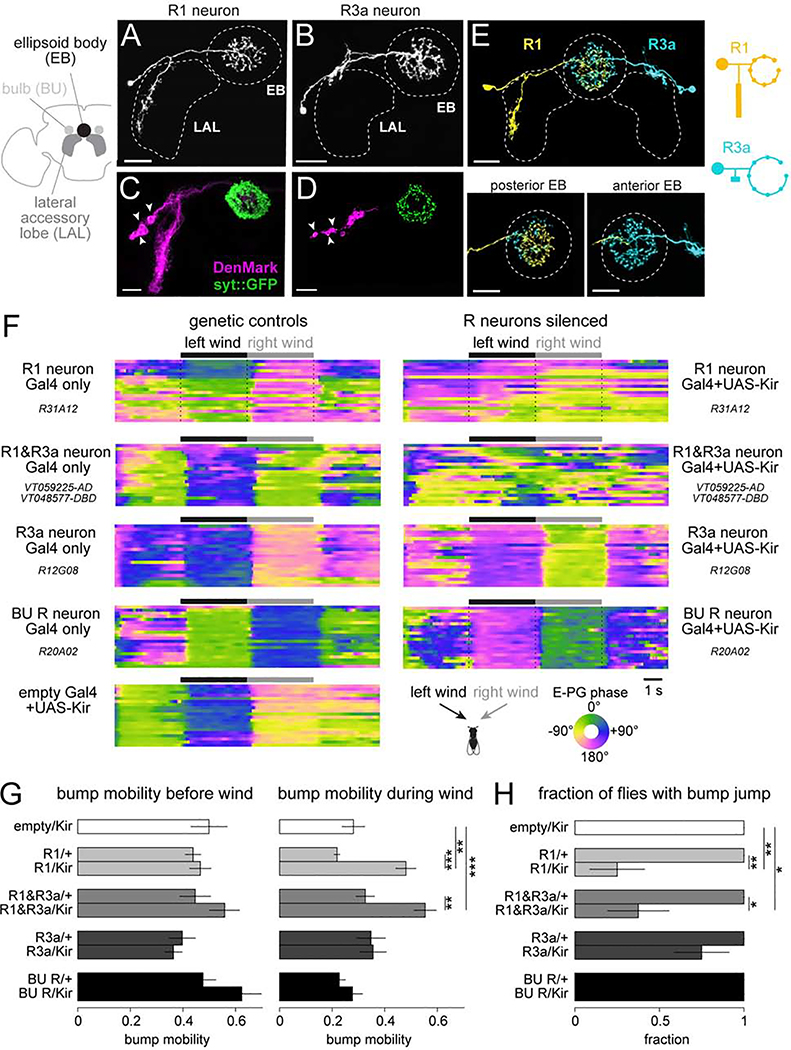
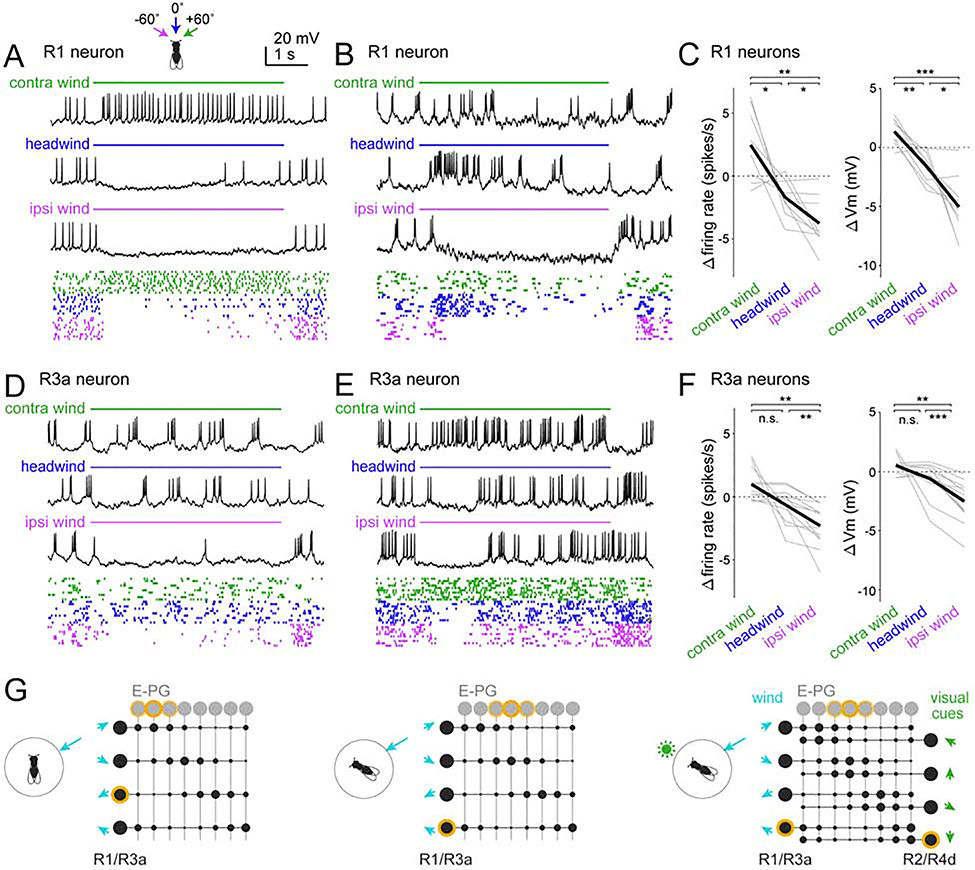
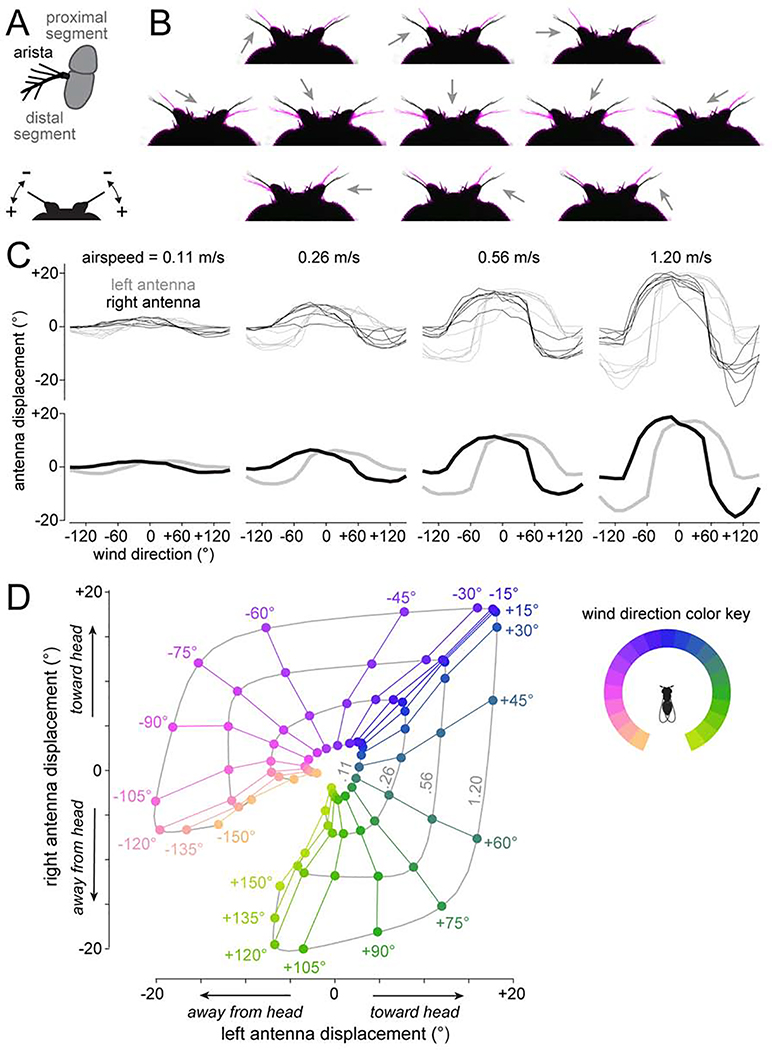
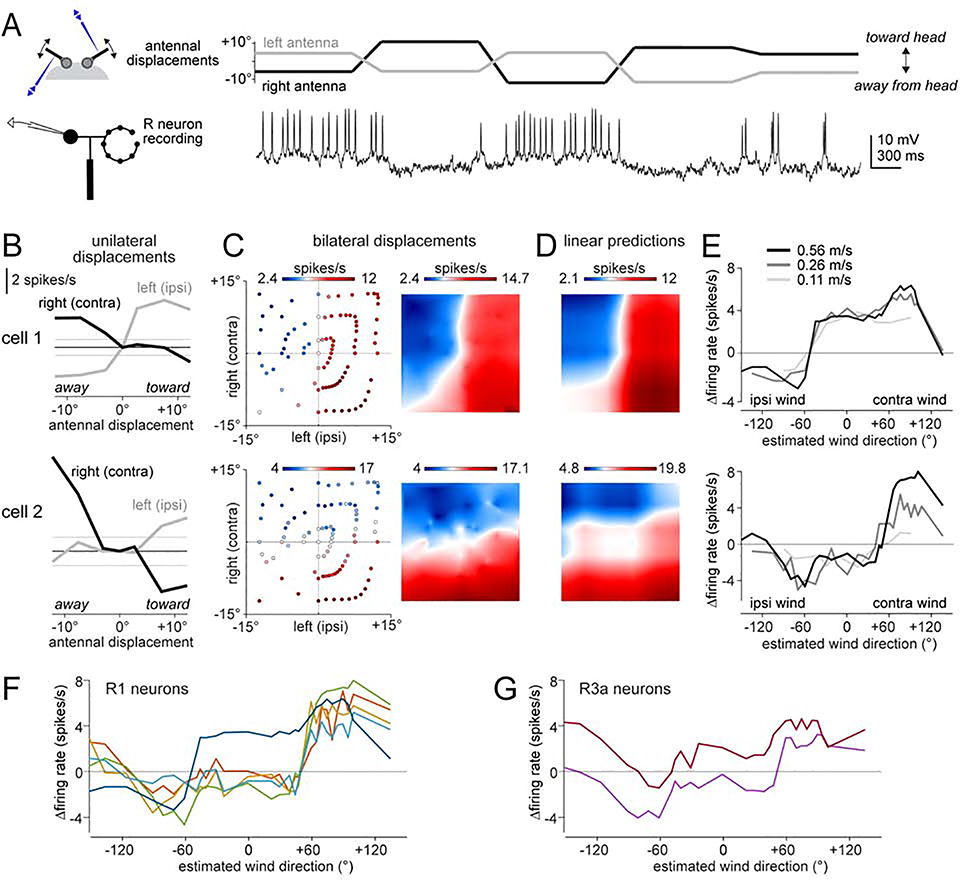
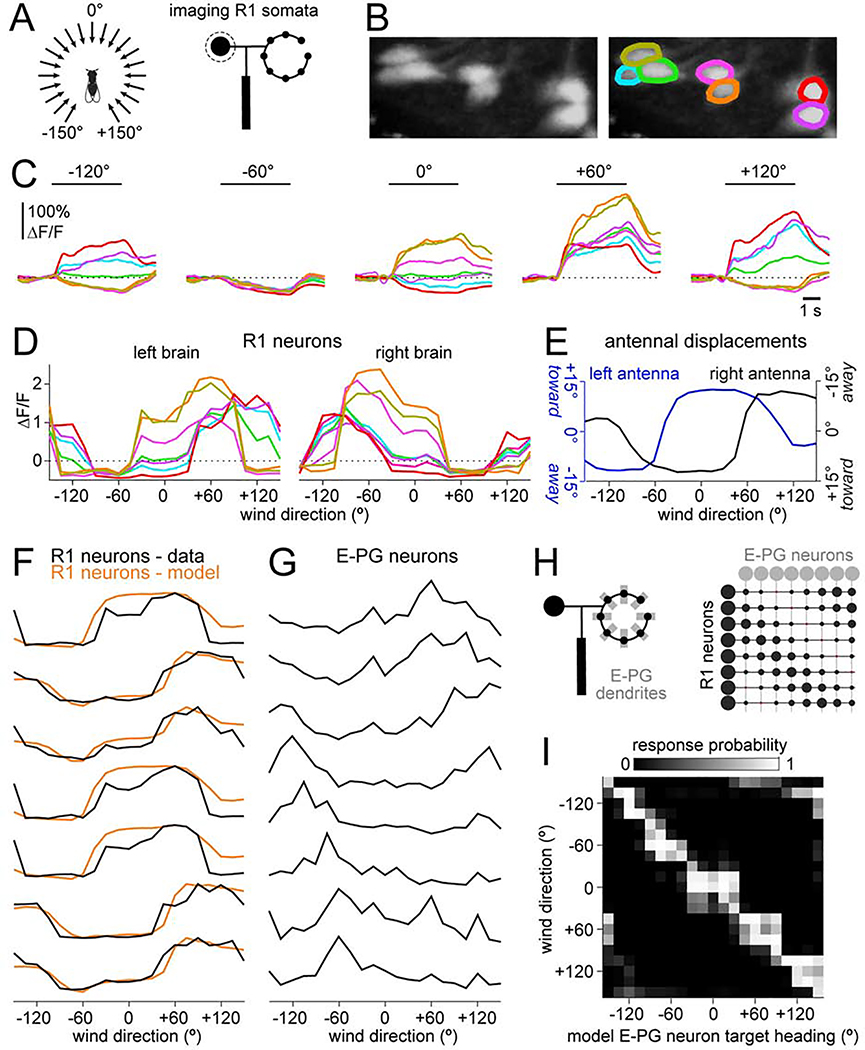
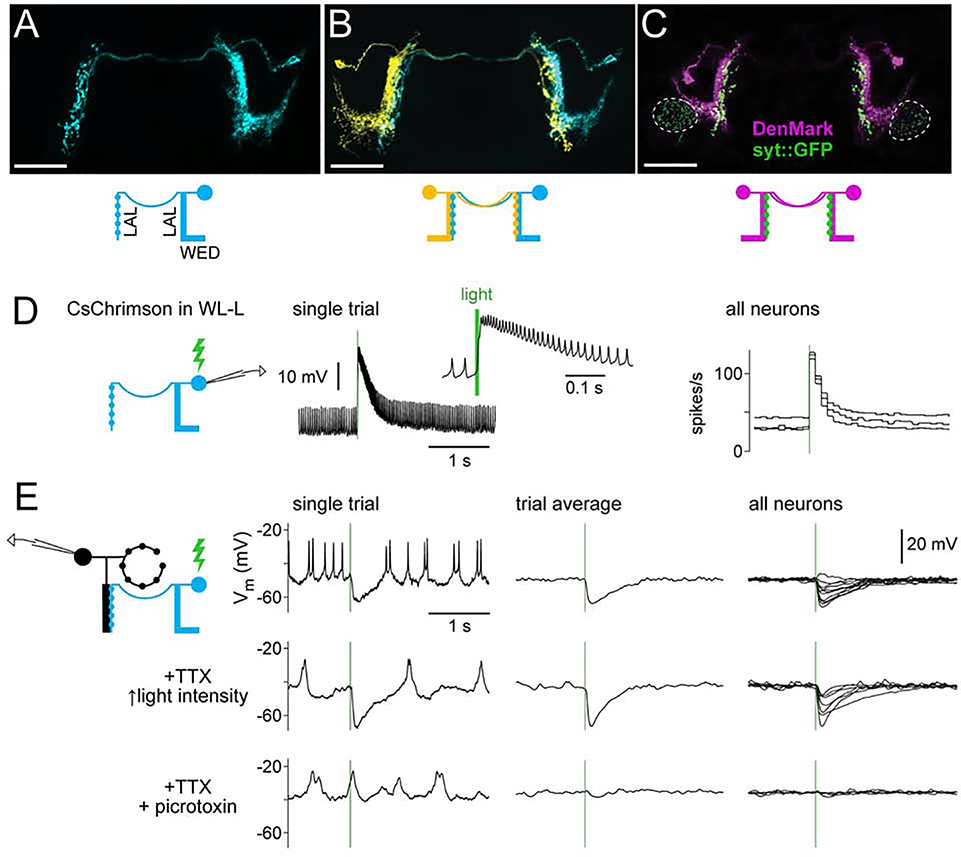
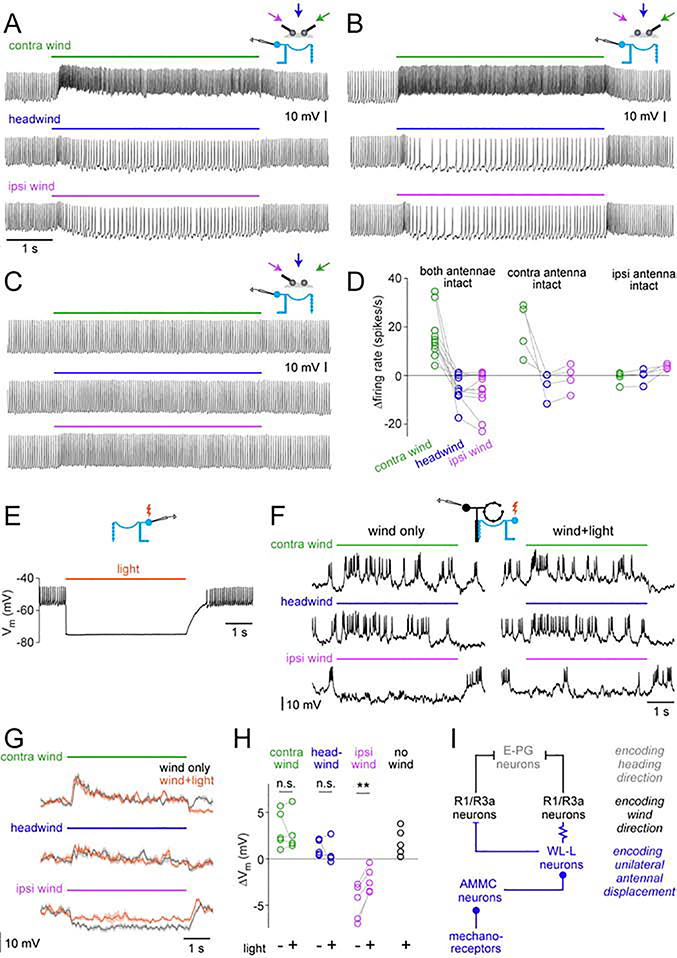
Spatial maps in the brain are most accurate when they are linked to external sensory cues. Here, we show that the compass in the Drosophila brain is linked to the direction of the wind. Shifting the wind rightward rotates the compass as if the fly were turning leftward, and vice versa. We describe the mechanisms of several computations that integrate wind information into the compass. First, an intensity-invariant representation of wind direction is computed by comparing left-right mechanosensory signals. Then, signals are reformatted to reduce the coding biases inherent in peripheral mechanics, and wind cues are brought into the same circular coordinate system that represents visual cues and self-motion signals. Because the compass incorporates both mechanosensory and visual cues, it should enable navigation under conditions where no single cue is consistently reliable. These results show how local sensory signals can be transformed into a global, multimodal, abstract representation of space.
Keywords: AMMC; Johnston’s organ; Ring neuron; central complex; ellipsoid body; lateral accessory lobe; mechanosensation; sensorimotor integration; wedge.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
Comment in
-
Insect Orientation: The Drosophila Wind Compass Pathway.Curr Biol. 2021 Jan 25;31(2):R83-R85. doi: 10.1016/j.cub.2020.11.033. Curr Biol. 2021. PMID: 33497638
References
-
- Amit DJ, Wong KYM, and Campbell C (1989). Perceptron learning with sign-constrained weights. J Phys A-Math Gen 22, 2039–2045.
-
- Baker DA, Beckingham KM, and Armstrong JD (2007). Functional dissection of the neural substrates for gravitaxic maze behavior in Drosophila melanogaster. J Comp Neurol 501, 756–764. - PubMed
-
- Bates AS, Franconville R, and Jefferis GSXE (2019a). neuprintr: R client utilities for interacting with the neuPrint connectome analysis service. R package version 0.4.0.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
