

E3 Ubiquitin Ligase Von Hippel-Lindau Protein Promotes Th17 Differentiation
- PMID: 32690659
- PMCID: PMC8167928
- DOI: 10.4049/jimmunol.2000243
E3 Ubiquitin Ligase Von Hippel-Lindau Protein Promotes Th17 Differentiation
Abstract
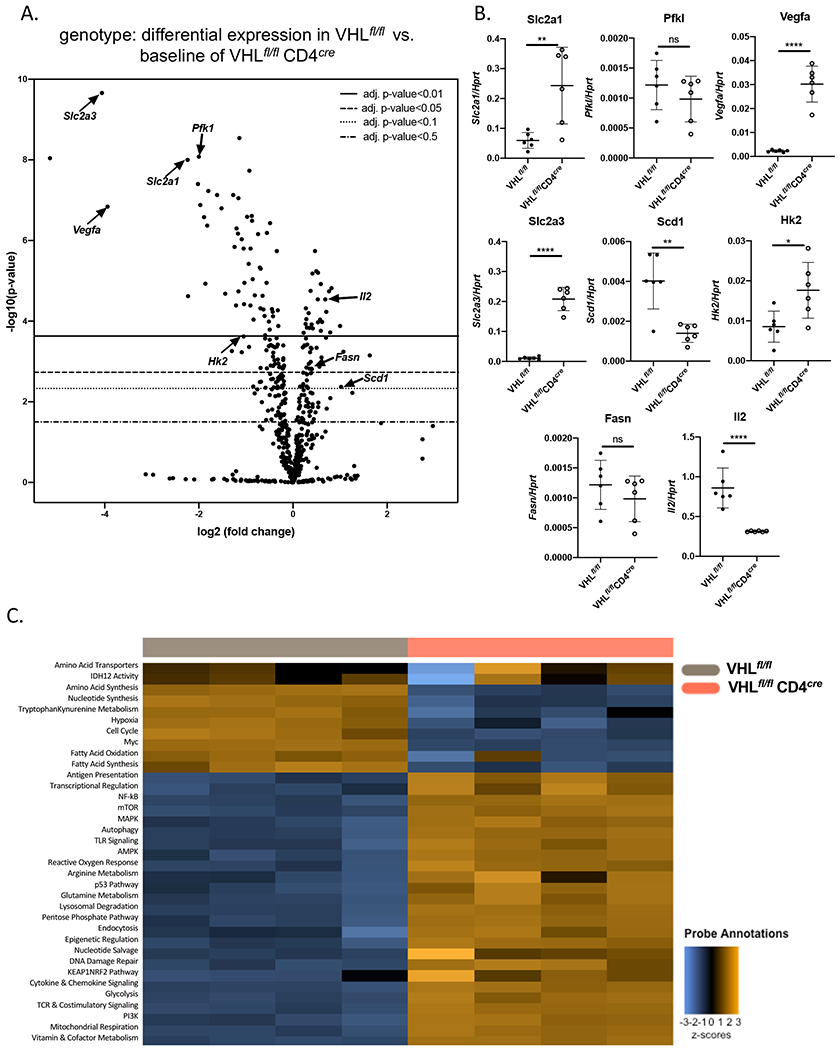
Von Hippel-Lindau (VHL) is an E3 ubiquitin ligase that targets proteins, including HIF-1α, for proteasomal degradation. VHL and HIF regulate the balance between glycolysis and oxidative phosphorylation, which is critical in highly dynamic T cells. HIF-1α positively regulates Th17 differentiation, a complex process in which quiescent naive CD4 T cells undergo transcriptional changes to effector cells, which are commonly dysregulated in autoimmune diseases. The role of VHL in Th17 cells is not known. In this study, we hypothesized VHL negatively regulates Th17 differentiation and deletion of VHL in CD4 T cells would elevate HIF-1α and increase Th17 differentiation. Unexpectedly, we found that VHL promotes Th17 differentiation. Mice deficient in VHL in their T cells were resistant to an autoimmune disease, experimental autoimmune encephalomyelitis, often mediated by Th17 cells. In vitro Th17 differentiation was impaired in VHL-deficient T cells. In the absence of VHL, Th17 cells had decreased activation of STAT3 and SMAD2, suggesting that VHL indirectly or directly regulates these critical signaling molecules. Gene expression analysis revealed that in Th17 cells, VHL regulates many cellular pathways, including genes encoding proteins involved indirectly or directly in the glycolysis pathway. Compared with wild-type, VHL-deficient Th17 cells had elevated glycolysis and glycolytic capacity. Our finding has implications on the design of therapeutics targeting the distinct metabolic needs of T cells to combat chronic inflammatory diseases.
Copyright © 2020 by The American Association of Immunologists, Inc.
Conflict of interest statement
Conflict of Interest
The authors have no conflicts of interest to report.
Figures







References
-
- Stockinger B, and Omenetti S. 2017. The dichotomous nature of T helper 17 cells. Nat Rev Immunol 17: 535–544. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
