

Monocyte-derived APCs are central to the response of PD1 checkpoint blockade and provide a therapeutic target for combination therapy
- PMID: 32690667
- PMCID: PMC7371367
- DOI: 10.1136/jitc-2020-000588
Monocyte-derived APCs are central to the response of PD1 checkpoint blockade and provide a therapeutic target for combination therapy
Abstract
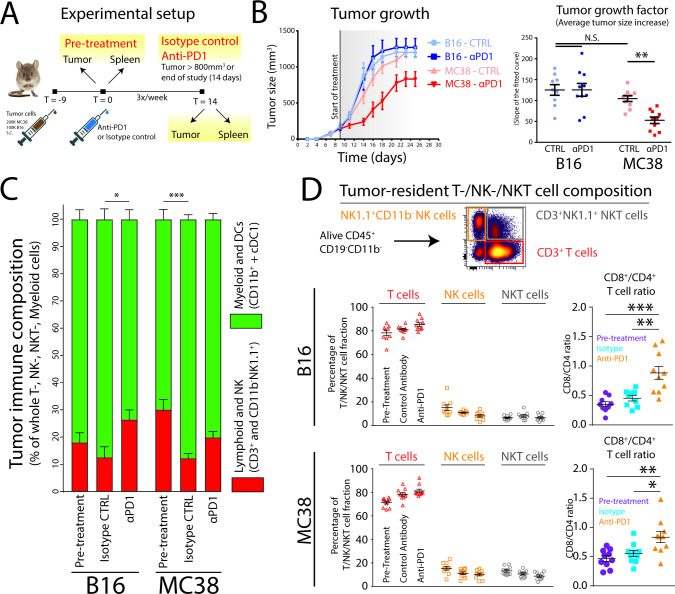
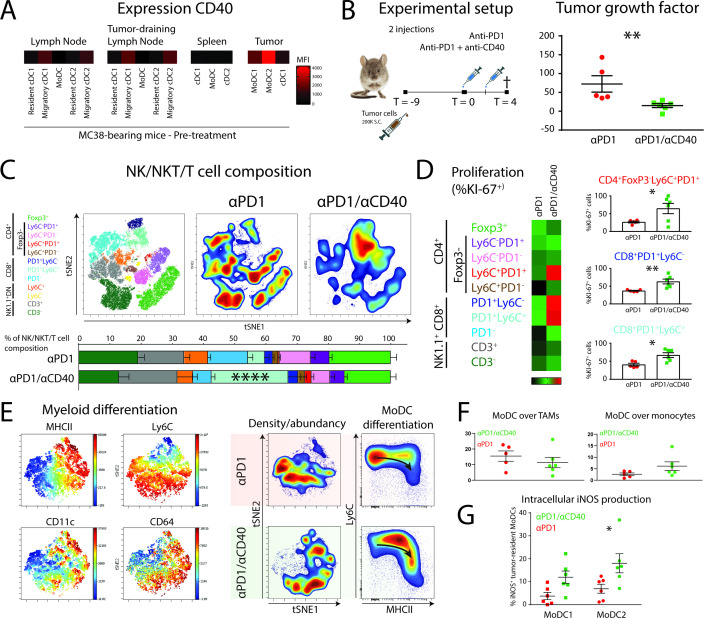
Background: PD1 immune checkpoint blockade (αPD1 ICB) has shown unparalleled success in treating many types of cancer. However, response to treatment does not always lead to tumor rejection. While αPD1 ICB relies on cytotoxic CD8+ T cells, antigen-presenting cells (APCs) at the tumor site are also needed for costimulation of tumor-infiltrating lymphocytes (TILs). It is still unclear how these APCs develop and function before and during αPD1 ICB or how they are associated with tumor rejection.
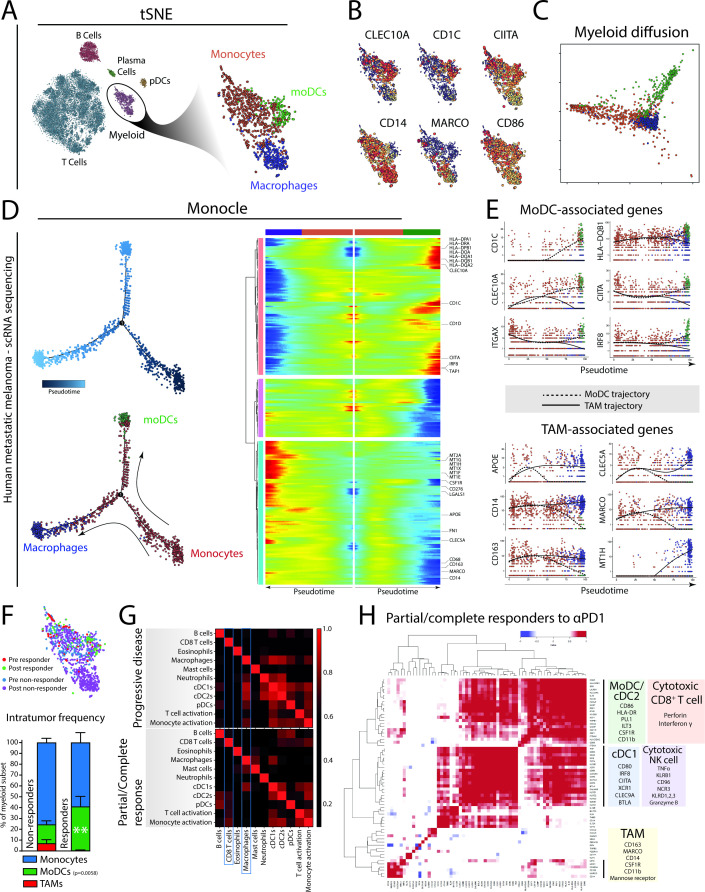
Methods: Here, we used B16 mouse melanoma and MC38 colorectal carcinoma tumor models, which show differential responses to αPD1 ICB. The immune composition of ICB insensitive B16 and sensitive MC38 were extensively investigated using multi-parameter flow cytometry and unsupervised clustering and trajectory analyses. We additionally analyzed existing single cell RNA sequencing data of the myeloid compartment of patients with melanoma undergoing αPD1 ICB. Lastly, we investigated the effect of CD40 agonistic antibody on the tumor-infiltrating monocyte-derived cells during αPD1 ICB.
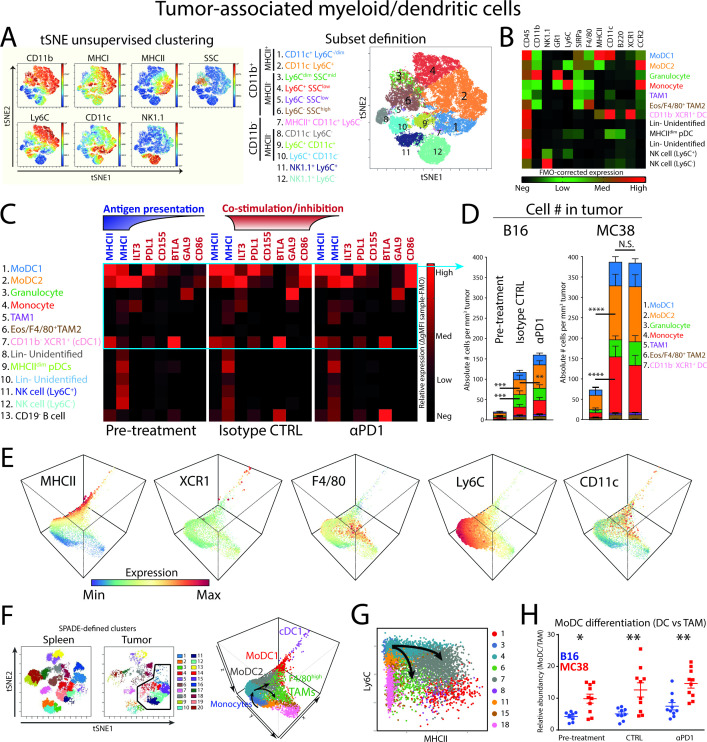
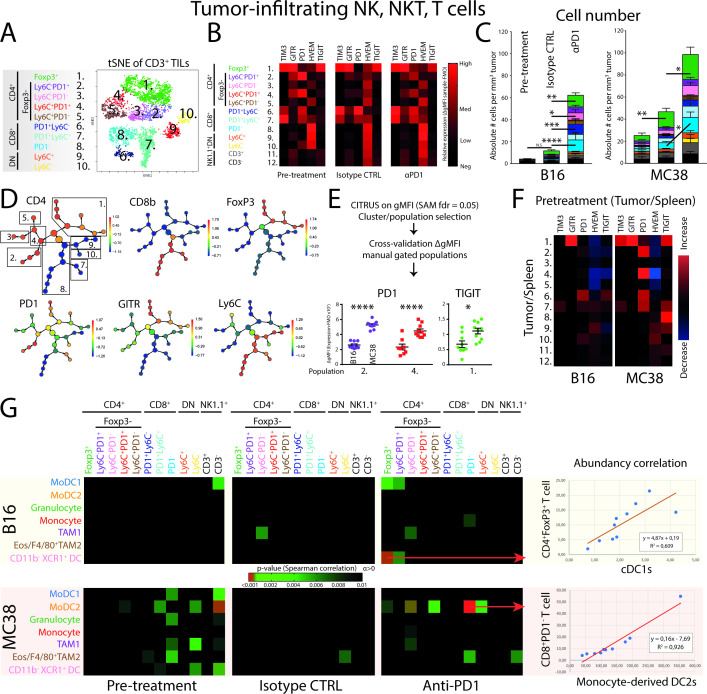
Results: We show that monocyte-derived dendritic cells (moDCs) express high levels of costimulatory molecules and are correlated with effector TILs in the tumor microenvironment (TME) after αPD1 ICB only in responding mouse tumor models. Tumor-resident moDCs showed distinct differentiation from monocytes in both mouse and human tumors. We further confirmed significant enrichment of tumor-resident differentiated moDCs in patients with melanoma responding to αPD1 ICB therapy compared with non-responding patients. Moreover, moDCs could be targeted by agonistic anti-CD40 antibody, supporting moDC differentiation, effector T-cell expansion and anti-tumor immunity.
Conclusion: The combined analysis of myeloid and lymphoid populations in the TME during successful and non-successful PD1 ICB led to the discovery of monocyte-to-DC differentiation linked to expanding T-cell populations. This differentiation was found in patients during ICB, which was significantly higher during successful ICB. The finding of tumor-infiltrating monocytes and differentiating moDCs as druggable target for rational combination therapy opens new avenues of anti-tumor therapy design.
Keywords: costimulatory and inhibitory T-cell receptors; dendritic cells; immunotherapy; programmed cell death 1 receptor; tumor microenvironment.
© Author(s) (or their employer(s)) 2020. Re-use permitted under CC BY. Published by BMJ.
Conflict of interest statement
Competing interests: None declared.
Figures
Similar articles
-
Anti-PD1 antibody enhances the anti-tumor efficacy of MUC1-MBP fusion protein vaccine via increasing Th1, Tc1 activity and decreasing the proportion of MDSC in the B16-MUC1 melanoma mouse model.Int Immunopharmacol. 2021 Dec;101(Pt A):108173. doi: 10.1016/j.intimp.2021.108173. Epub 2021 Oct 1. Int Immunopharmacol. 2021. PMID: 34607233
-
TIGIT and PD-1 Immune Checkpoint Pathways Are Associated With Patient Outcome and Anti-Tumor Immunity in Glioblastoma.Front Immunol. 2021 May 7;12:637146. doi: 10.3389/fimmu.2021.637146. eCollection 2021. Front Immunol. 2021. PMID: 34025646 Free PMC article.
-
Tumor CD155 Expression Is Associated with Resistance to Anti-PD1 Immunotherapy in Metastatic Melanoma.Clin Cancer Res. 2020 Jul 15;26(14):3671-3681. doi: 10.1158/1078-0432.CCR-19-3925. Epub 2020 Apr 28. Clin Cancer Res. 2020. PMID: 32345648
-
Antigen presenting cells in cancer immunity and mediation of immune checkpoint blockade.Clin Exp Metastasis. 2024 Aug;41(4):333-349. doi: 10.1007/s10585-023-10257-z. Epub 2024 Jan 23. Clin Exp Metastasis. 2024. PMID: 38261139 Free PMC article. Review.
-
Combination immunotherapy strategies for glioblastoma.J Neurooncol. 2021 Feb;151(3):375-391. doi: 10.1007/s11060-020-03481-0. Epub 2021 Feb 21. J Neurooncol. 2021. PMID: 33611705 Review.
Cited by
-
Predictive circulating biomarkers of the response to anti-PD-1 immunotherapy in advanced HER2 negative breast cancer.Clin Transl Med. 2025 Mar;15(3):e70255. doi: 10.1002/ctm2.70255. Clin Transl Med. 2025. PMID: 40000397 Free PMC article.
-
Characteristics of a CCL21 Gene-Modified Dendritic Cell Vaccine Utilized for a Clinical Trial in Non-Small Cell Lung Cancer.Mol Cancer Ther. 2025 Feb 4;24(2):286-298. doi: 10.1158/1535-7163.MCT-24-0435. Mol Cancer Ther. 2025. PMID: 39559833 Free PMC article. Clinical Trial.
-
Blood-based molecular and cellular biomarkers of early response to neoadjuvant PD-1 blockade in patients with non-small cell lung cancer.Cancer Cell Int. 2024 Jun 29;24(1):225. doi: 10.1186/s12935-024-03412-3. Cancer Cell Int. 2024. PMID: 38951894 Free PMC article.
-
CD33 Expression on Peripheral Blood Monocytes Predicts Efficacy of Anti-PD-1 Immunotherapy Against Non-Small Cell Lung Cancer.Front Immunol. 2022 Apr 14;13:842653. doi: 10.3389/fimmu.2022.842653. eCollection 2022. Front Immunol. 2022. PMID: 35493454 Free PMC article.
-
Cellular mechanisms of combining innate immunity activation with PD-1/PD-L1 blockade in treatment of colorectal cancer.Mol Cancer. 2024 Nov 12;23(1):252. doi: 10.1186/s12943-024-02166-w. Mol Cancer. 2024. PMID: 39529058 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous