

TNNT2 mutations in the tropomyosin binding region of TNT1 disrupt its role in contractile inhibition and stimulate cardiac dysfunction
- PMID: 32690703
- PMCID: PMC7414051
- DOI: 10.1073/pnas.2001692117
TNNT2 mutations in the tropomyosin binding region of TNT1 disrupt its role in contractile inhibition and stimulate cardiac dysfunction
Abstract
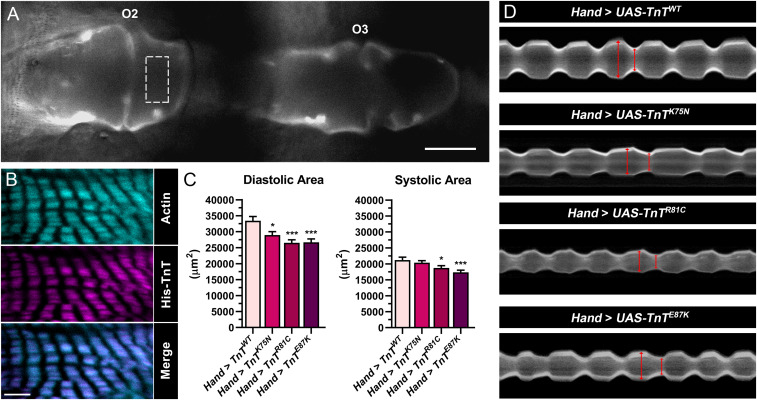
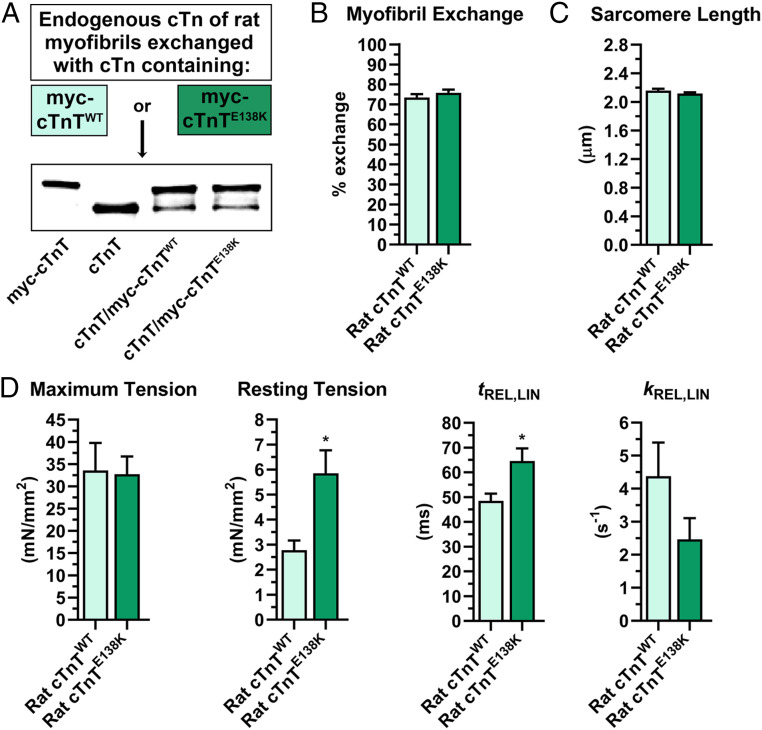
Muscle contraction is regulated by the movement of end-to-end-linked troponin-tropomyosin complexes over the thin filament surface, which uncovers or blocks myosin binding sites along F-actin. The N-terminal half of troponin T (TnT), TNT1, independently promotes tropomyosin-based, steric inhibition of acto-myosin associations, in vitro. Recent structural models additionally suggest TNT1 may restrain the uniform, regulatory translocation of tropomyosin. Therefore, TnT potentially contributes to striated muscle relaxation; however, the in vivo functional relevance and molecular basis of this noncanonical role remain unclear. Impaired relaxation is a hallmark of hypertrophic and restrictive cardiomyopathies (HCM and RCM). Investigating the effects of cardiomyopathy-causing mutations could help clarify TNT1's enigmatic inhibitory property. We tested the hypothesis that coupling of TNT1 with tropomyosin's end-to-end overlap region helps anchor tropomyosin to an inhibitory position on F-actin, where it deters myosin binding at rest, and that, correspondingly, cross-bridge cycling is defectively suppressed under diastolic/low Ca2+ conditions in the presence of HCM/RCM lesions. The impact of TNT1 mutations on Drosophila cardiac performance, rat myofibrillar and cardiomyocyte properties, and human TNT1's propensity to inhibit myosin-driven, F-actin-tropomyosin motility were evaluated. Our data collectively demonstrate that removing conserved, charged residues in TNT1's tropomyosin-binding domain impairs TnT's contribution to inhibitory tropomyosin positioning and relaxation. Thus, TNT1 may modulate acto-myosin activity by optimizing F-actin-tropomyosin interfacial contacts and by binding to actin, which restrict tropomyosin's movement to activating configurations. HCM/RCM mutations, therefore, highlight TNT1's essential role in contractile regulation by diminishing its tropomyosin-anchoring effects, potentially serving as the initial trigger of pathology in our animal models and humans.
Keywords: Drosophila; cardiomyopathy; diastolic dysfunction; tropomyosin; troponin T.
Conflict of interest statement
The authors declare no competing interest.
Figures




References
-
- Tobacman L. S., Thin filament-mediated regulation of cardiac contraction. Annu. Rev. Physiol. 58, 447–481 (1996). - PubMed
-
- Geeves M. A., “Thin filament regulation” in Comprehensive Biophysics, Egelman E. H., Goldman Y. E., Ostap E. M., Eds. (Academic, Oxford, United Kingdom, 2012), Vol. 4, pp. 251–267.
-
- Lehman W., Thin filament structure and the steric blocking model. Compr. Physiol. 6, 1043–1069 (2016). - PubMed
-
- Hitchcock-DeGregori S. E., Barua B., “Tropomyosin structure, function, and interactions: A dynamic regulator” in Fibrous Proteins: Structures and Mechanisms, Parry D. A. D., Squire J., Eds. (Springer, 2017), pp. 253–284. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
