

Allosteric deactivation of PIFs and EIN3 by microproteins in light control of plant development
- PMID: 32694206
- PMCID: PMC7414296
- DOI: 10.1073/pnas.2002313117
Allosteric deactivation of PIFs and EIN3 by microproteins in light control of plant development
Abstract
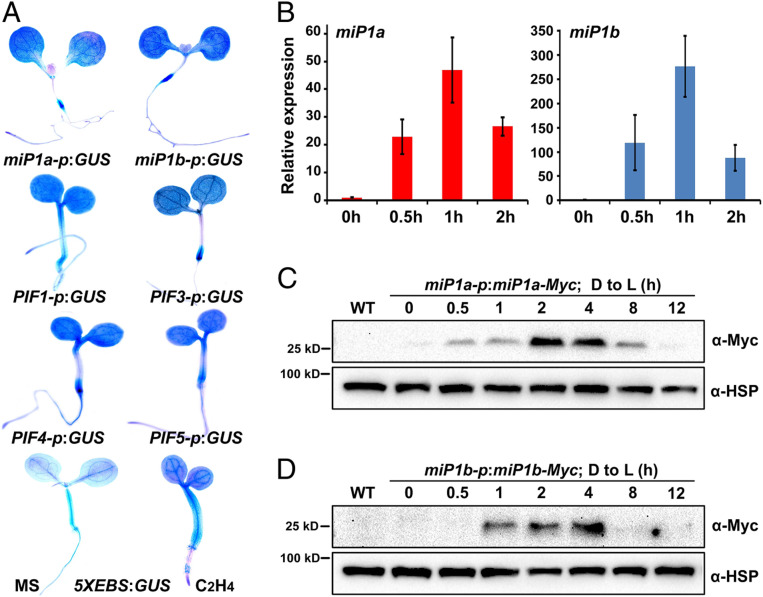
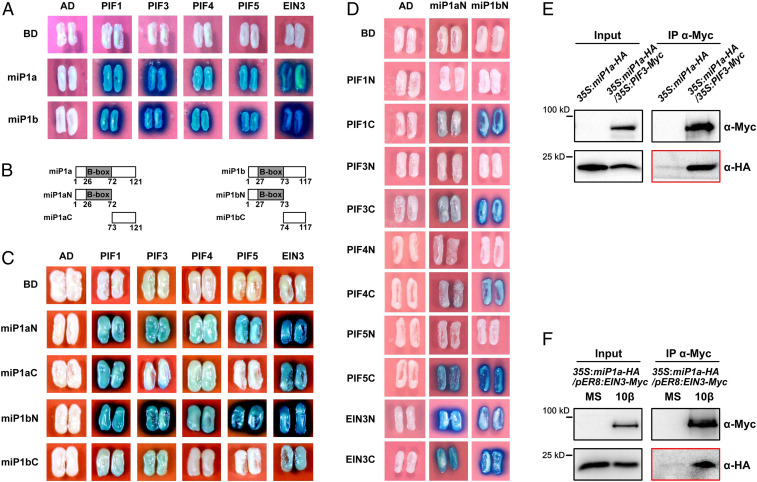
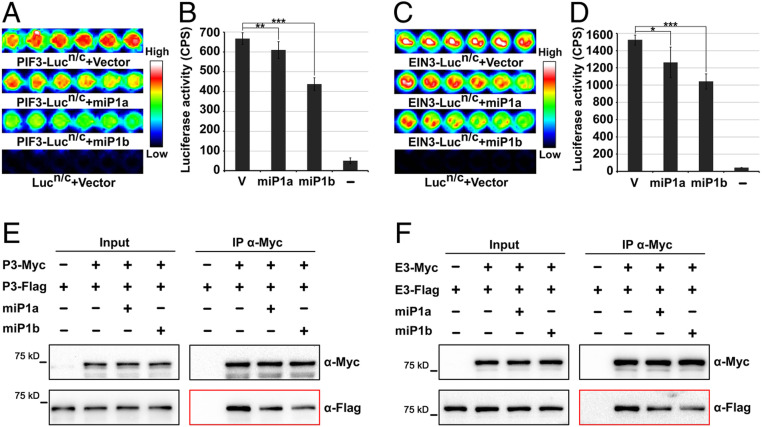
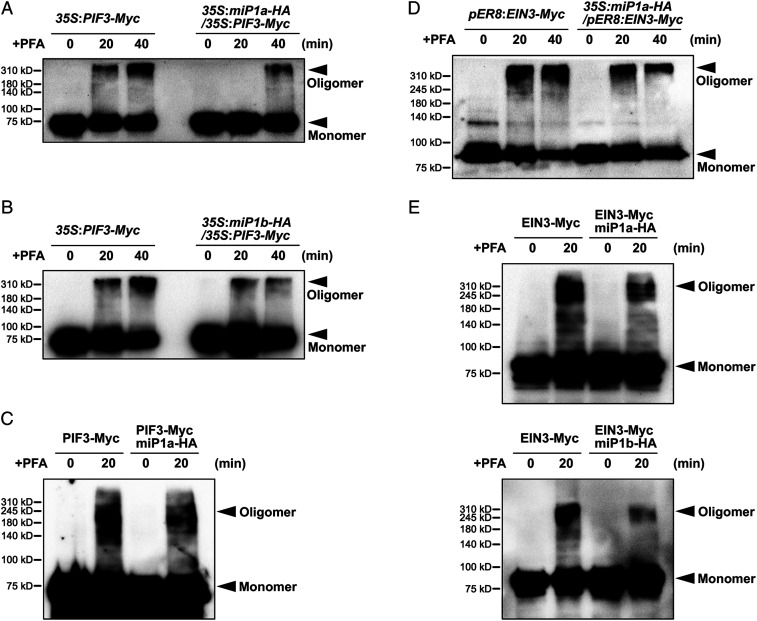
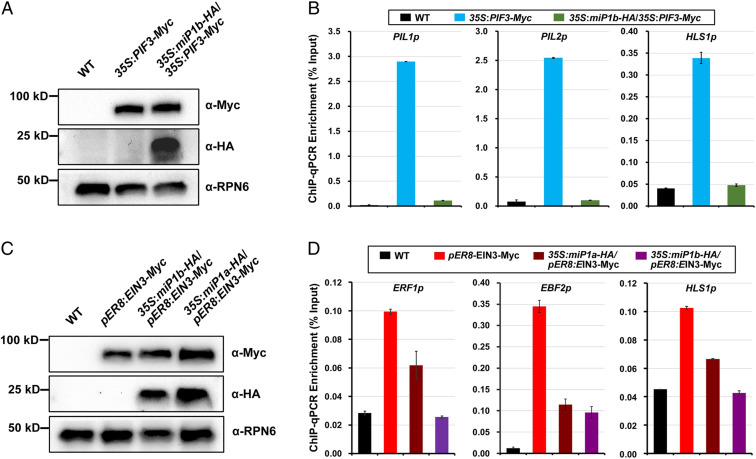
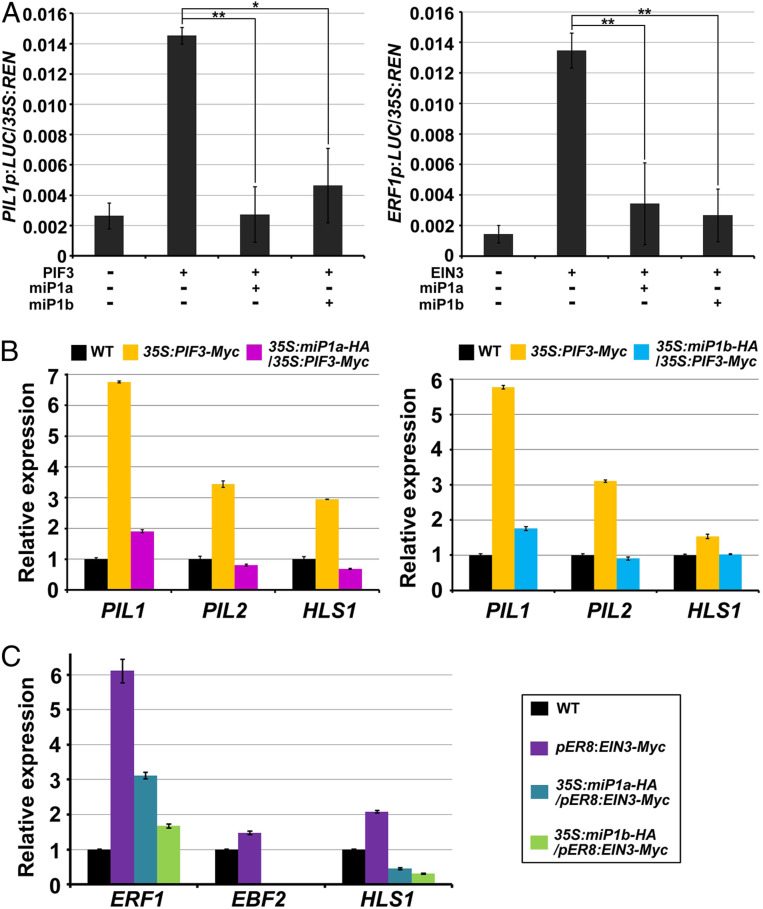
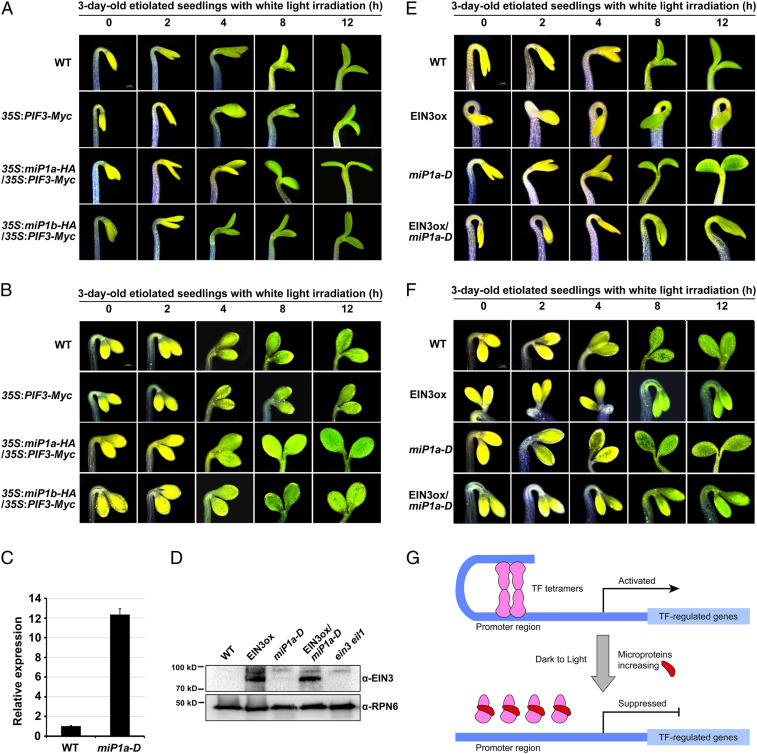
Buried seedlings undergo dramatic developmental transitions when they emerge from soil into sunlight. As central transcription factors suppressing light responses, PHYTOCHROME-INTERACTING FACTORs (PIFs) and ETHYLENE-INSENSITIVE 3 (EIN3) actively function in darkness and must be promptly repressed upon light to initiate deetiolation. Microproteins are evolutionarily conserved small single-domain proteins that act as posttranslational regulators in eukaryotes. Although hundreds to thousands of microproteins are predicted to exist in plants, their target molecules, biological roles, and mechanisms of action remain largely unknown. Here, we show that two microproteins, miP1a and miP1b (miP1a/b), are robustly stimulated in the dark-to-light transition. miP1a/b are primarily expressed in cotyledons and hypocotyl, exhibiting tissue-specific patterns similar to those of PIFs and EIN3 We demonstrate that PIFs and EIN3 assemble functional oligomers by self-interaction, while miP1a/b directly interact with and disrupt the oligomerization of PIFs and EIN3 by forming nonfunctional protein complexes. As a result, the DNA binding capacity and transcriptional activity of PIFs and EIN3 are predominantly suppressed. These biochemical findings are further supported by genetic evidence. miP1a/b positively regulate photomorphogenic development, and constitutively expressing miP1a/b rescues the delayed apical hook unfolding and cotyledon development of plants overexpressing PIFs and EIN3 Our study reveals that microproteins provide a temporal and negative control of the master transcription factors' oligomerization to achieve timely developmental transitions upon environmental changes.
Keywords: EIN3/EIL1; PIFs; light signaling; microprotein; protein oligomerization.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
