

Insulin signaling and glucose metabolism in different hepatoma cell lines deviate from hepatocyte physiology toward a convergent aberrant phenotype
- PMID: 32694512
- PMCID: PMC7374613
- DOI: 10.1038/s41598-020-68721-9
Insulin signaling and glucose metabolism in different hepatoma cell lines deviate from hepatocyte physiology toward a convergent aberrant phenotype
Abstract
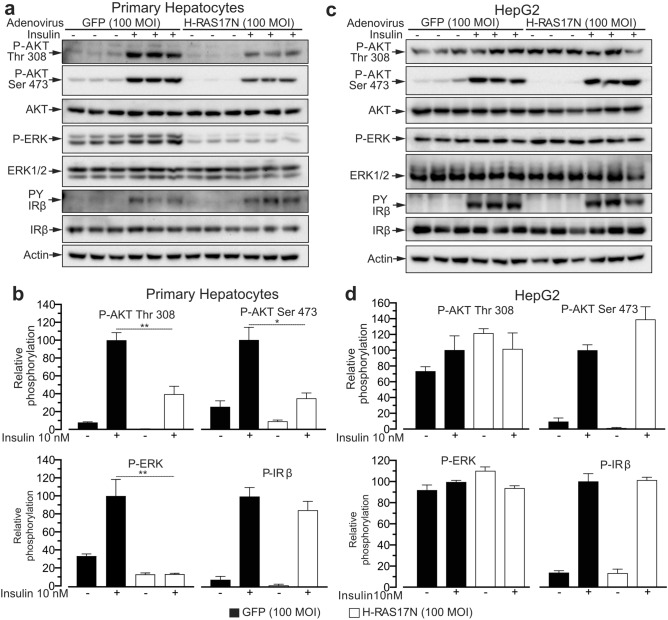
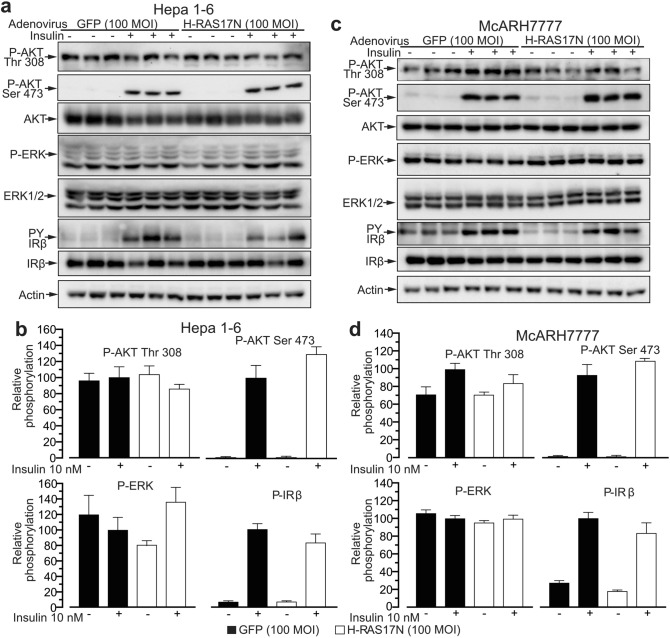
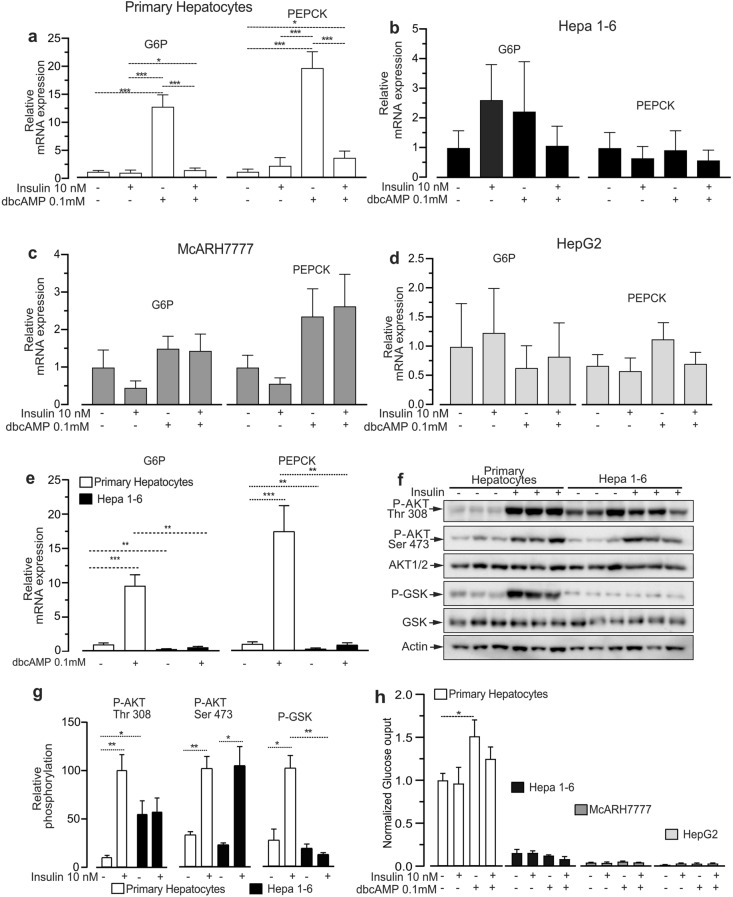
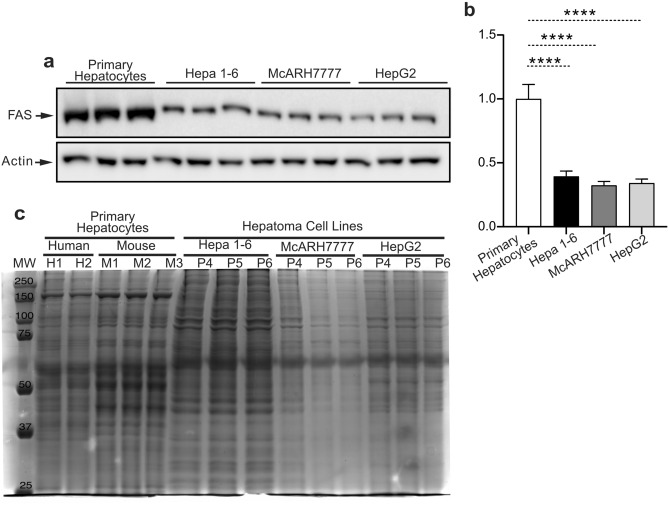
Hepatoma cell lines are widely used to model the hepatocyte for insulin signaling and fatty liver disease. However, a direct comparison of insulin action in primary hepatocytes and in hepatoma cell lines is needed to validate this model and to better understand liver cancer. Here we have investigated insulin signaling, gluconeogenic gene expression, glucose production, and fatty acid synthase abundance in primary hepatocytes and in HepG2, Hepa 1-6, and McARH7777 hepatoma cell lines. Differences in the electrophoretic profiles of protein extracts from human and mouse primary hepatocytes and the hepatoma cells lines are shown. Compared to primary hepatocytes, hepatoma cells showed high basal phosphorylation of AKT at Thr 308 and constitutively activated RAS-MAPK signaling, which were resistant to the dominant negative Ras mutant H-Ras17N. Hepatoma cell lines also showed defective expression of gluconeogenic enzymes, insulin unresponsive GSK phosphorylation, and marginal glucose production. Hepatoma cells also showed lower protein levels of fatty acid synthase and a largely distinct protein electrophoresis profile from hepatocytes but similar between different hepatoma lines. We conclude that hepatoma cell lines do not accurately model the hepatocyte for insulin action but may be valuable tools to investigate the proteomic changes conferring to hepatocellular carcinoma its peculiar metabolisms.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
