Integrating the inputs that shape pancreatic islet hormone release
- PMID: 32694675
- PMCID: PMC7378277
- DOI: 10.1038/s42255-019-0148-2
Integrating the inputs that shape pancreatic islet hormone release
Abstract
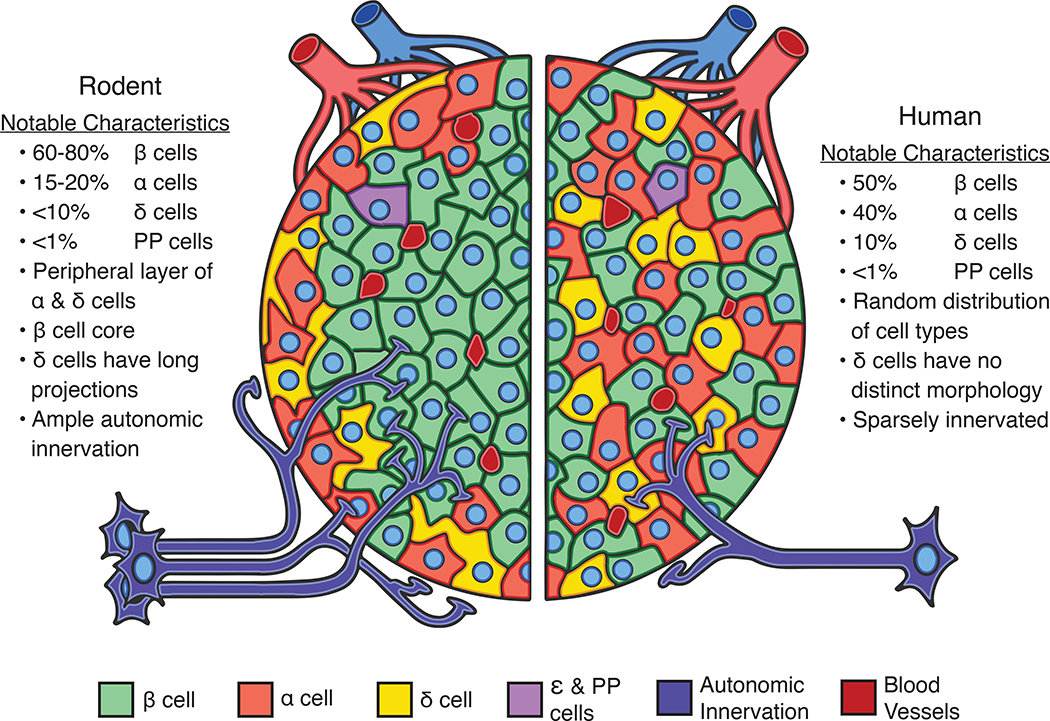
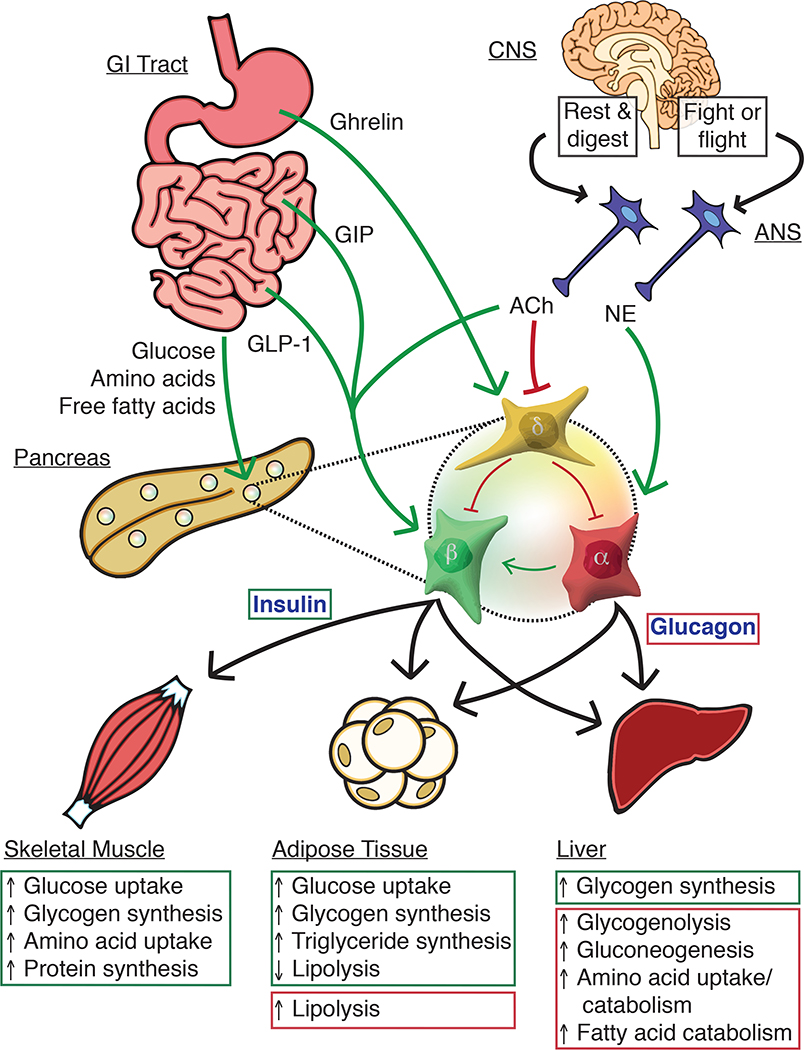
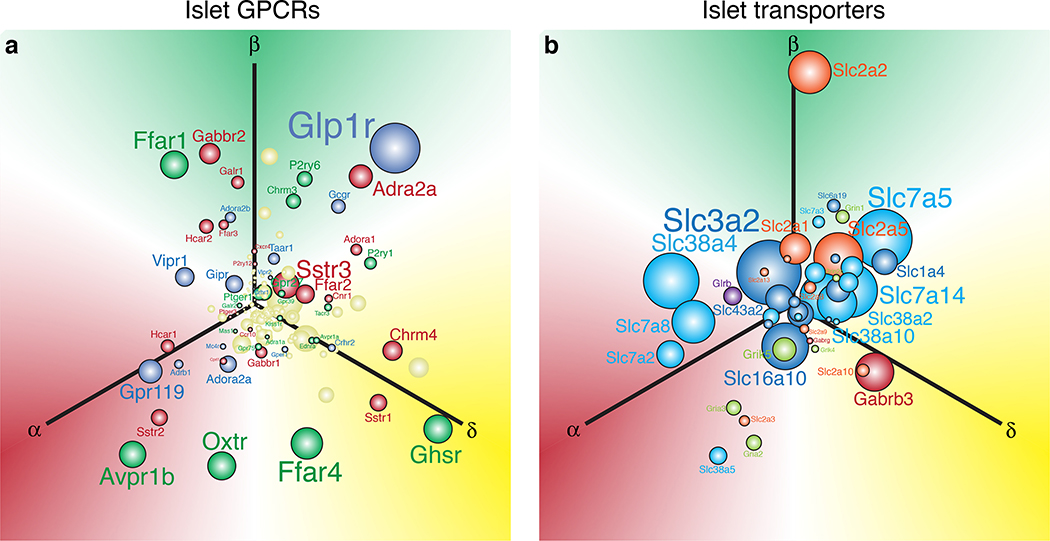
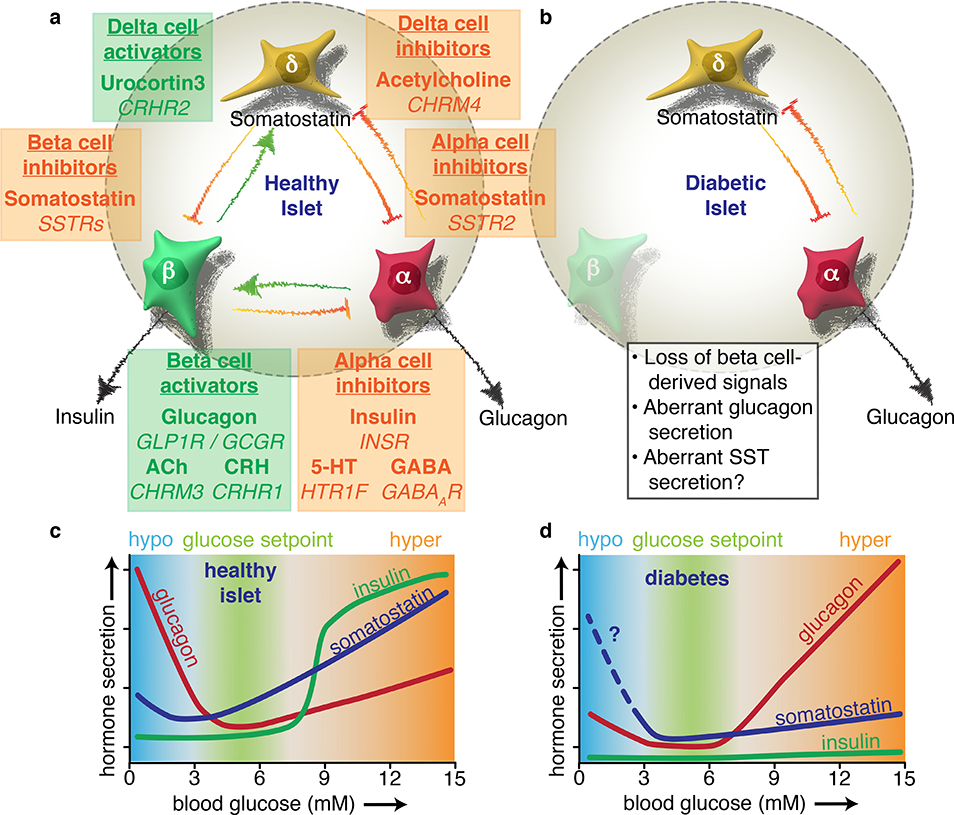
The pancreatic islet is a complex mini organ composed of a variety of endocrine cells and their support cells, which together tightly control blood glucose homeostasis. Changes in glucose concentration are commonly regarded as the chief signal controlling insulin-secreting beta cells, glucagon-secreting alpha cells and somatostatin-secreting delta cells. However, each of these cell types is highly responsive to a multitude of endocrine, paracrine, nutritional and neural inputs, which collectively shape the final endocrine output of the islet. Here, we review the principal inputs for each islet-cell type and the physiological circumstances in which these signals arise, through the prism of the insights generated by the transcriptomes of each of the major endocrine-cell types. A comprehensive integration of the factors that influence blood glucose homeostasis is essential to successfully improve therapeutic strategies for better diabetes management.
Figures