

Neurogenesis From Embryo to Adult - Lessons From Flies and Mice
- PMID: 32695783
- PMCID: PMC7339912
- DOI: 10.3389/fcell.2020.00533
Neurogenesis From Embryo to Adult - Lessons From Flies and Mice
Erratum in
-
Corrigendum: Neurogenesis From Embryo to Adult - Lessons From Flies and Mice.Front Cell Dev Biol. 2020 Aug 13;8:686. doi: 10.3389/fcell.2020.00686. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32903900 Free PMC article.
Abstract
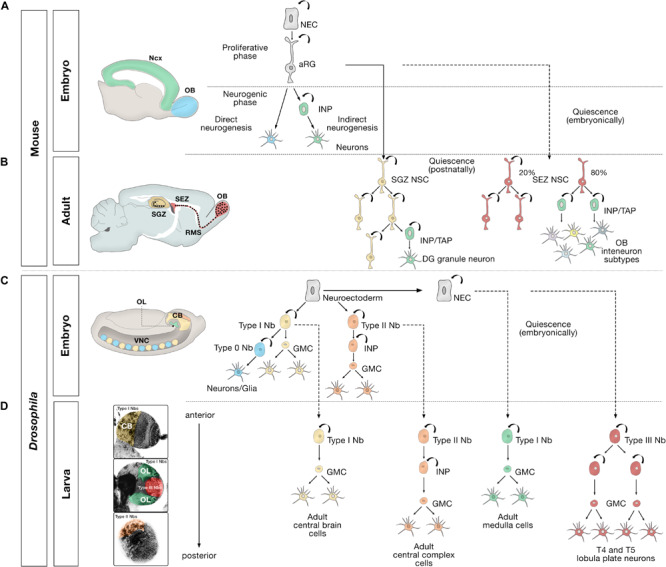
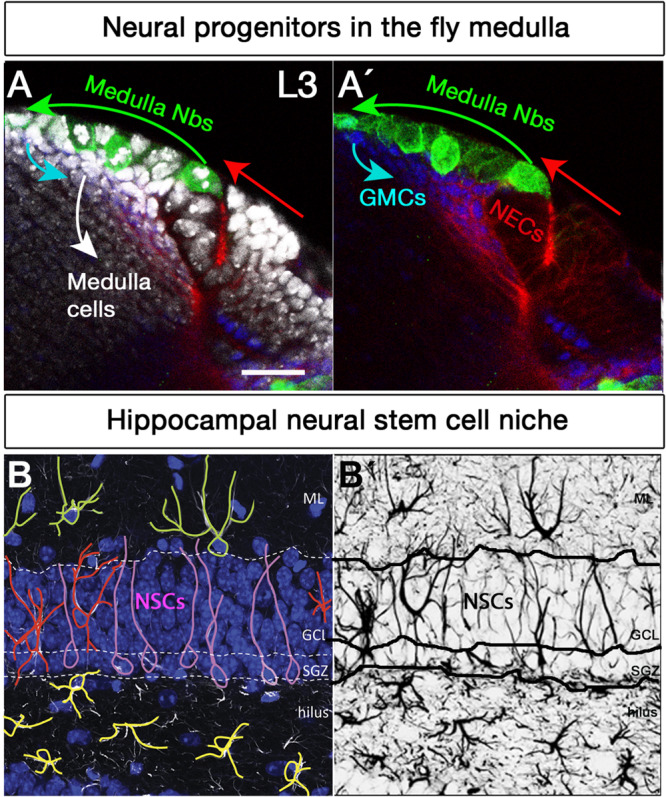
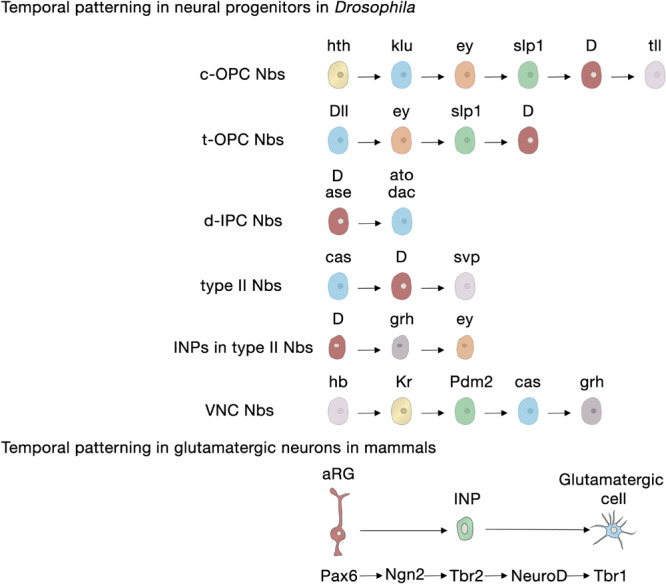
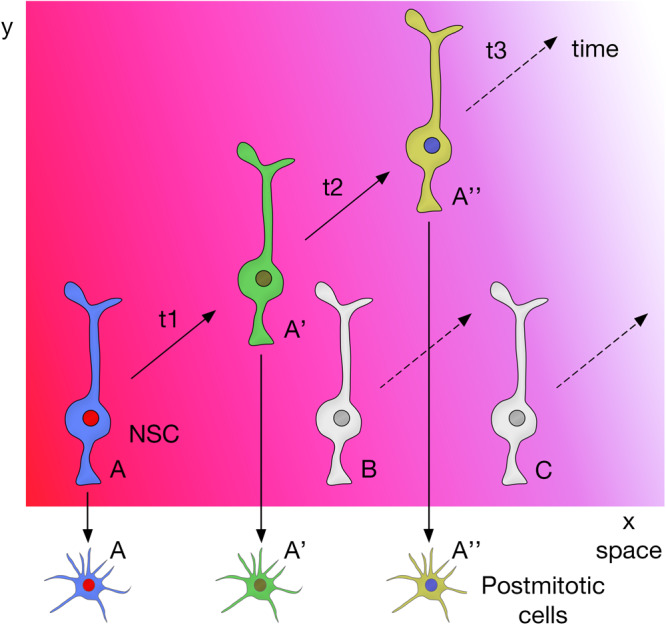
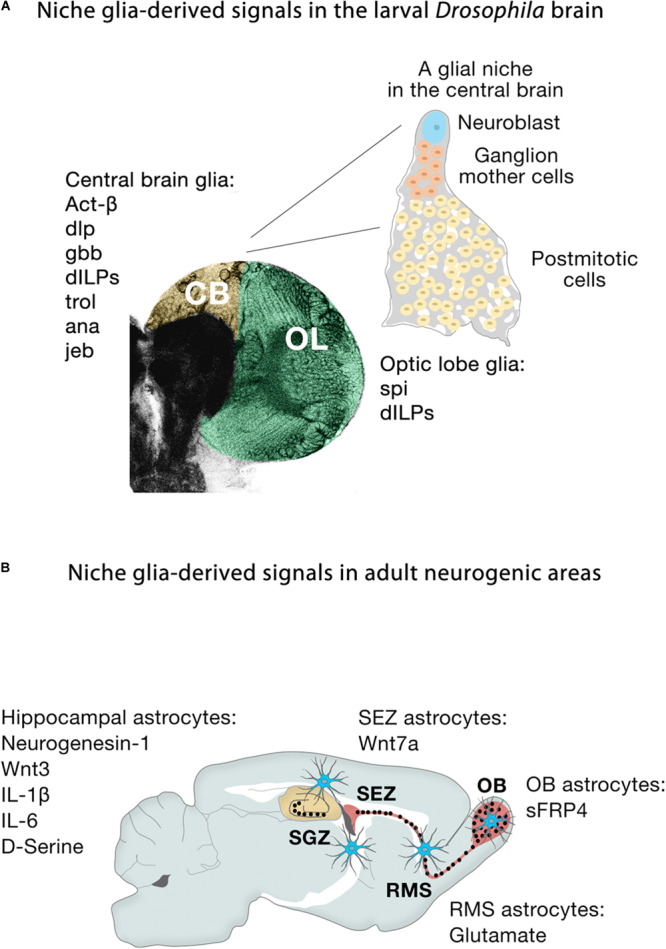
The human brain is composed of billions of cells, including neurons and glia, with an undetermined number of subtypes. During the embryonic and early postnatal stages, the vast majority of these cells are generated from neural progenitors and stem cells located in all regions of the neural tube. A smaller number of neurons will continue to be generated throughout our lives, in localized neurogenic zones, mainly confined at least in rodents to the subependymal zone of the lateral ventricles and the subgranular zone of the hippocampal dentate gyrus. During neurogenesis, a combination of extrinsic cues interacting with temporal and regional intrinsic programs are thought to be critical for increasing neuronal diversity, but their underlying mechanisms need further elucidation. In this review, we discuss the recent findings in Drosophila and mammals on the types of cell division and cell interactions used by neural progenitors and stem cells to sustain neurogenesis, and how they are influenced by glia.
Keywords: adult neurogenesis; extrinsic factors; glia; intrinsic factors; neural progenitors; neural stem cells; neurogenesis; niche.
Copyright © 2020 Mira and Morante.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases