

Elevated Glucose Levels Favor SARS-CoV-2 Infection and Monocyte Response through a HIF-1α/Glycolysis-Dependent Axis
- PMID: 32697943
- PMCID: PMC7367032
- DOI: 10.1016/j.cmet.2020.07.007
Elevated Glucose Levels Favor SARS-CoV-2 Infection and Monocyte Response through a HIF-1α/Glycolysis-Dependent Axis
Erratum in
-
Elevated Glucose Levels Favor SARS-CoV-2 Infection and Monocyte Response through a HIF-1α/Glycolysis-Dependent Axis.Cell Metab. 2020 Sep 1;32(3):498-499. doi: 10.1016/j.cmet.2020.07.015. Cell Metab. 2020. PMID: 32877692 Free PMC article. No abstract available.
Abstract
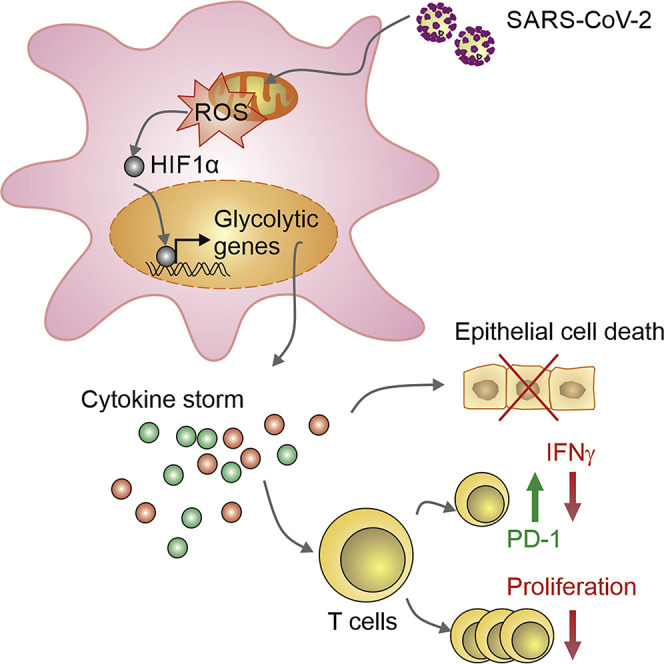
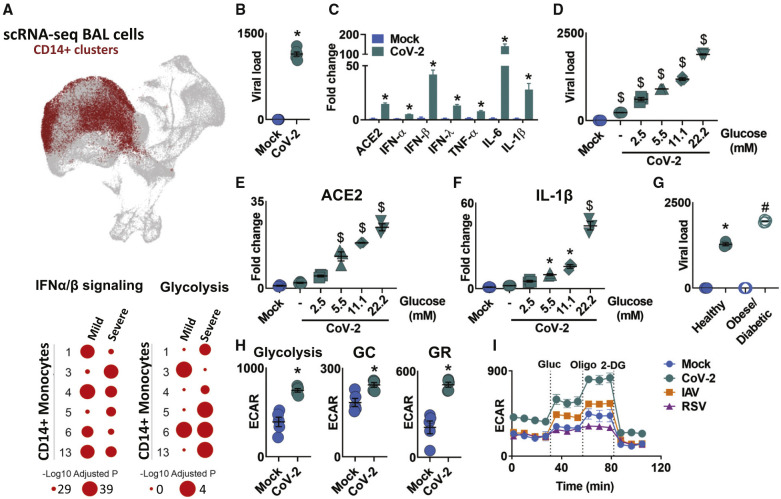
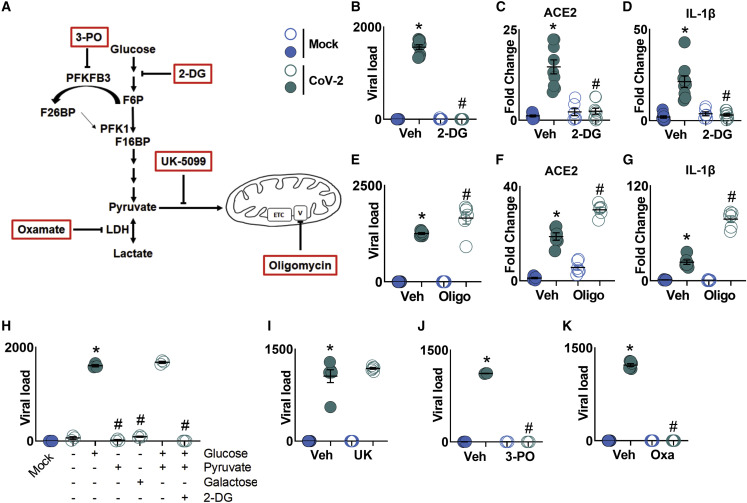
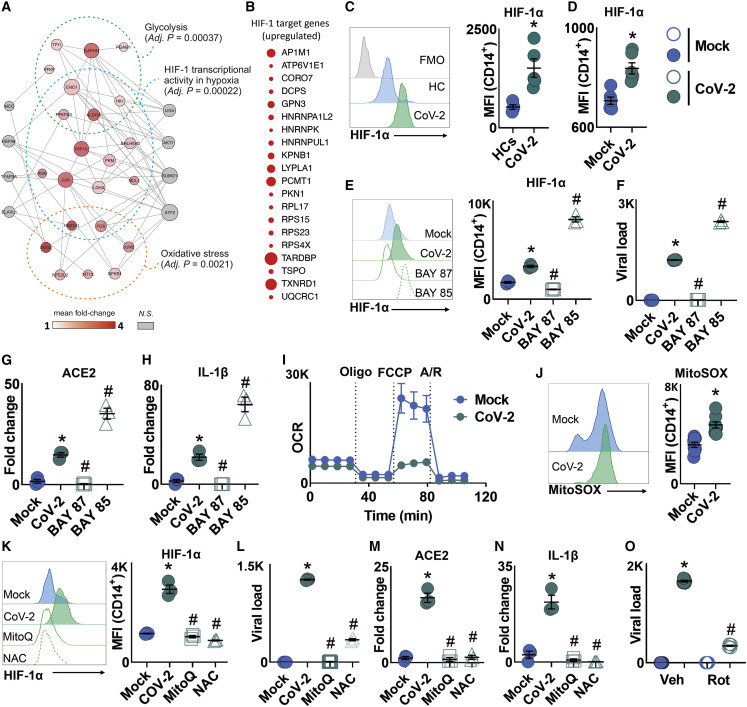
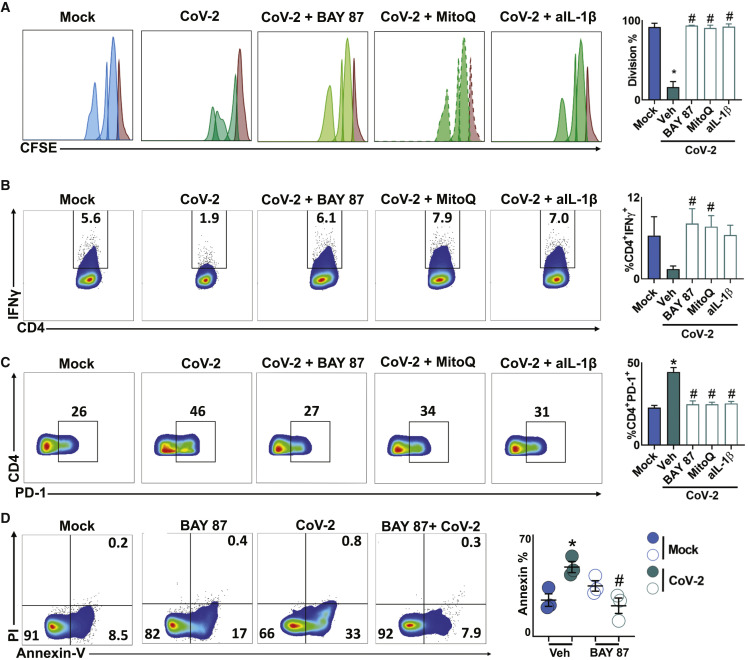
COVID-19 can result in severe lung injury. It remained to be determined why diabetic individuals with uncontrolled glucose levels are more prone to develop the severe form of COVID-19. The molecular mechanism underlying SARS-CoV-2 infection and what determines the onset of the cytokine storm found in severe COVID-19 patients are unknown. Monocytes and macrophages are the most enriched immune cell types in the lungs of COVID-19 patients and appear to have a central role in the pathogenicity of the disease. These cells adapt their metabolism upon infection and become highly glycolytic, which facilitates SARS-CoV-2 replication. The infection triggers mitochondrial ROS production, which induces stabilization of hypoxia-inducible factor-1α (HIF-1α) and consequently promotes glycolysis. HIF-1α-induced changes in monocyte metabolism by SARS-CoV-2 infection directly inhibit T cell response and reduce epithelial cell survival. Targeting HIF-1ɑ may have great therapeutic potential for the development of novel drugs to treat COVID-19.
Keywords: Covid-19; HIF-1alpha; SARS-CoV-2; diabetes; glycolysis; inflammation; interferon; metabolism; mitochondria; monocyte.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests Authors declare no competing interests.
Figures
Comment in
-
SARS-CoV-2 has a sweet tooth.Nat Rev Immunol. 2020 Aug;20(8):460. doi: 10.1038/s41577-020-0368-4. Nat Rev Immunol. 2020. PMID: 32533110 Free PMC article. No abstract available.
-
Targeting glucose metabolism for treatment of COVID-19.Signal Transduct Target Ther. 2021 Mar 6;6(1):112. doi: 10.1038/s41392-021-00532-4. Signal Transduct Target Ther. 2021. PMID: 33677470 Free PMC article. No abstract available.
References
-
- Baardman J., Verberk S.G.S., Prange K.H.M., van Weeghel M., van der Velden S., Ryan D.G., Wüst R.C.I., Neele A.E., Speijer D., Denis S.W. A defective pentose phosphate pathway reduces inflammatory macrophage responses during hypercholesterolemia. Cell Rep. 2018;25:2044–2052.e5. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
