

METTL1-mediated m7G methylation maintains pluripotency in human stem cells and limits mesoderm differentiation and vascular development
- PMID: 32698871
- PMCID: PMC7374972
- DOI: 10.1186/s13287-020-01814-4
METTL1-mediated m7G methylation maintains pluripotency in human stem cells and limits mesoderm differentiation and vascular development
Abstract
Background: 7-Methylguanosine (m7G) is one of the most conserved modifications in nucleosides within tRNAs and rRNAs. It plays essential roles in the regulation of mRNA export, splicing, and translation. Recent studies highlighted the importance of METTL1-mediated m7G tRNA methylome in the self-renewal of mouse embryonic stem cells (mESCs) through its ability to regulate mRNA translation. However, the exact mechanisms by which METTL1 regulates pluripotency and differentiation in human induced pluripotent stem cells (hiPSCs) remain unknown. In this study, we evaluated the functions and underlying molecular mechanisms of METTL1 in regulating hiPSC self-renewal and differentiation in vivo and in vitro.
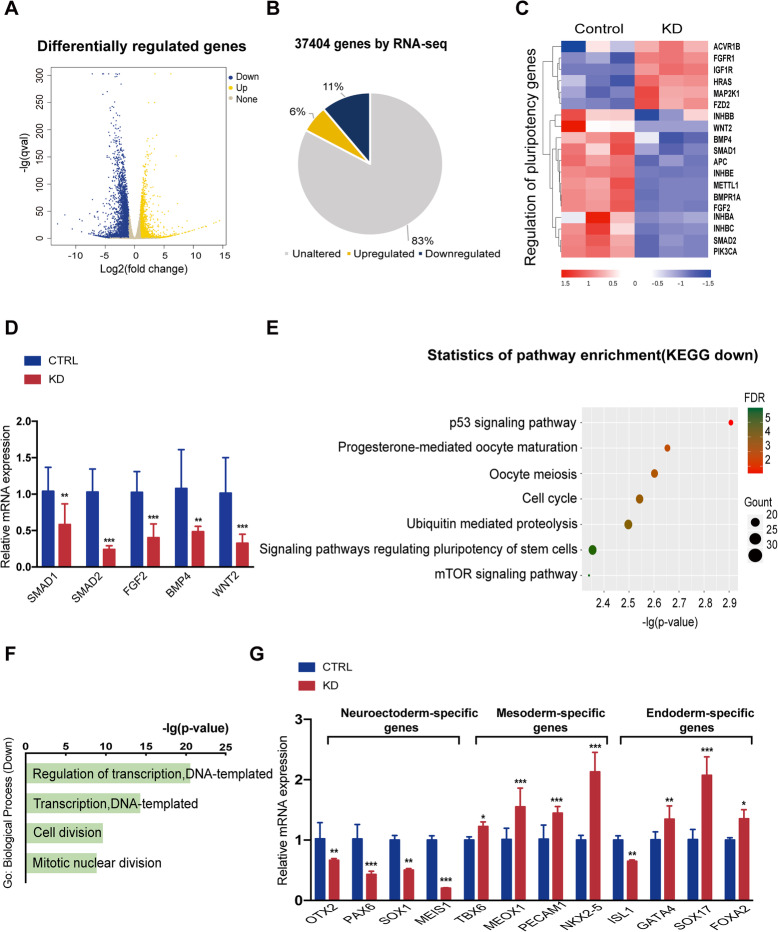
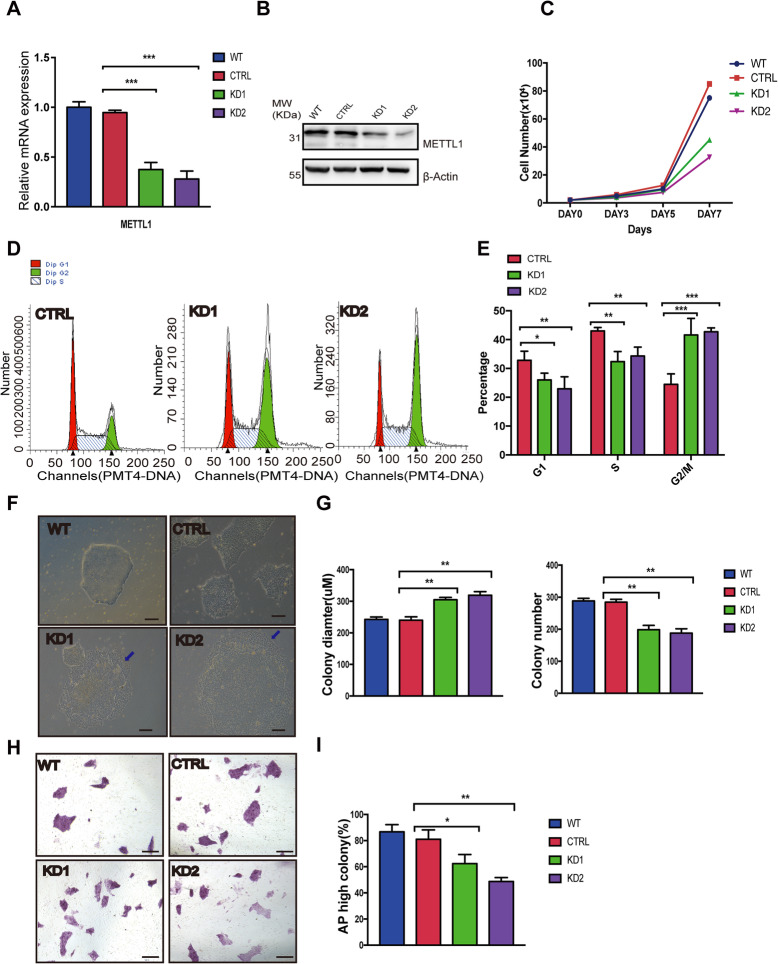
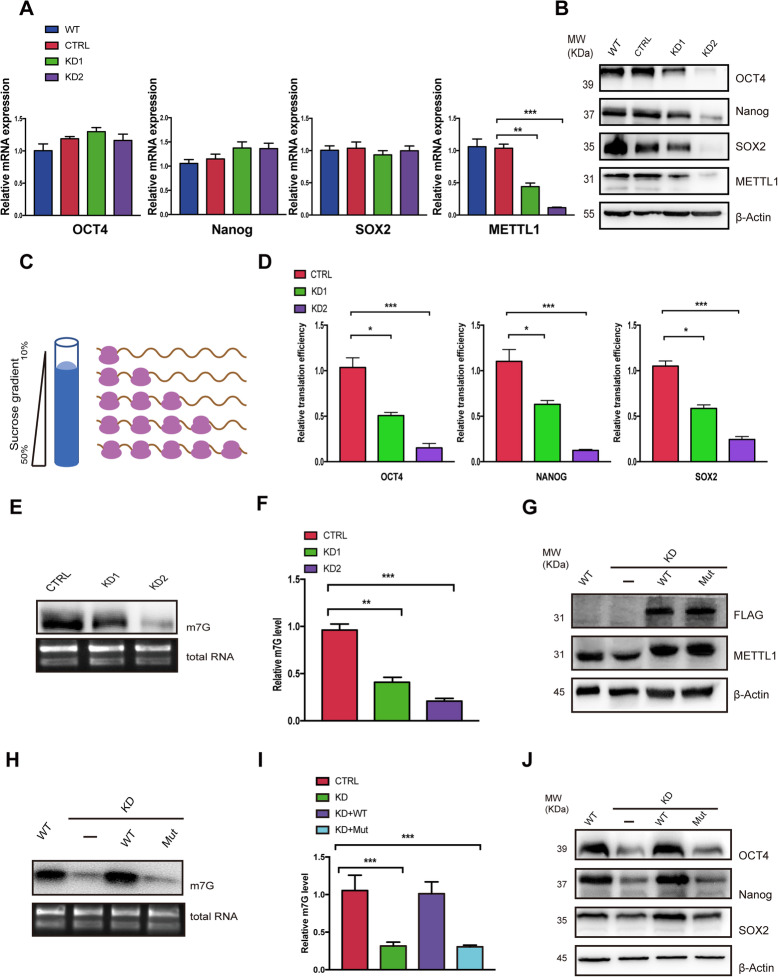
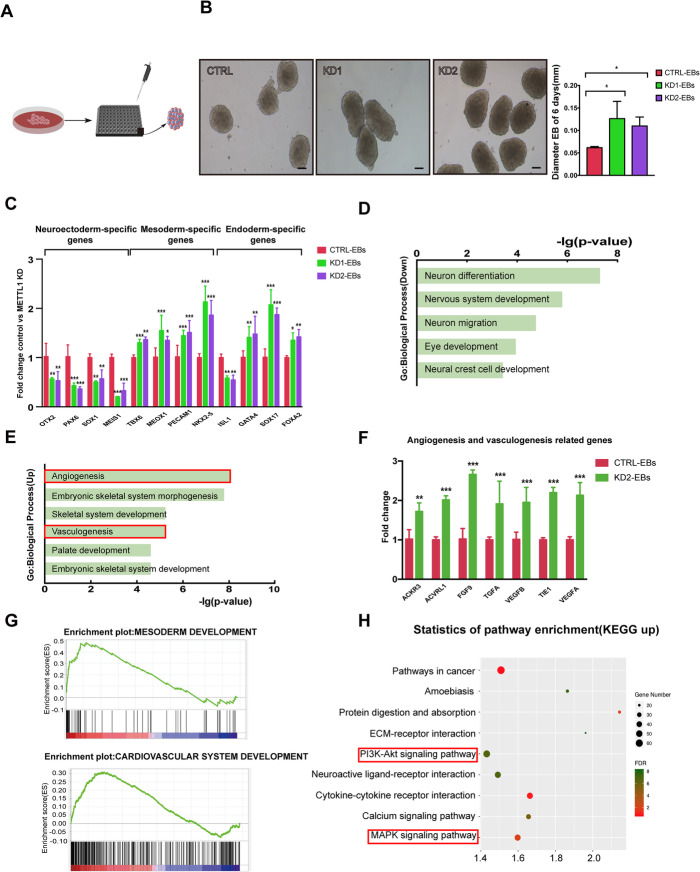
Methods: By establishing METTL1 knockdown (KD) hiPSCs, gene expression profiling was performed by RNA sequencing followed by pathway analyses. Anti-m7G northwestern assay was used to identify m7G modifications in tRNAs and mRNAs. Polysome profiling was used to assess the translation efficiency of the major pluripotent transcription factors. Moreover, the in vitro and in vivo differentiation capacities of METTL1-KD hiPSCs were assessed in embryoid body (EB) formation and teratoma formation assays.
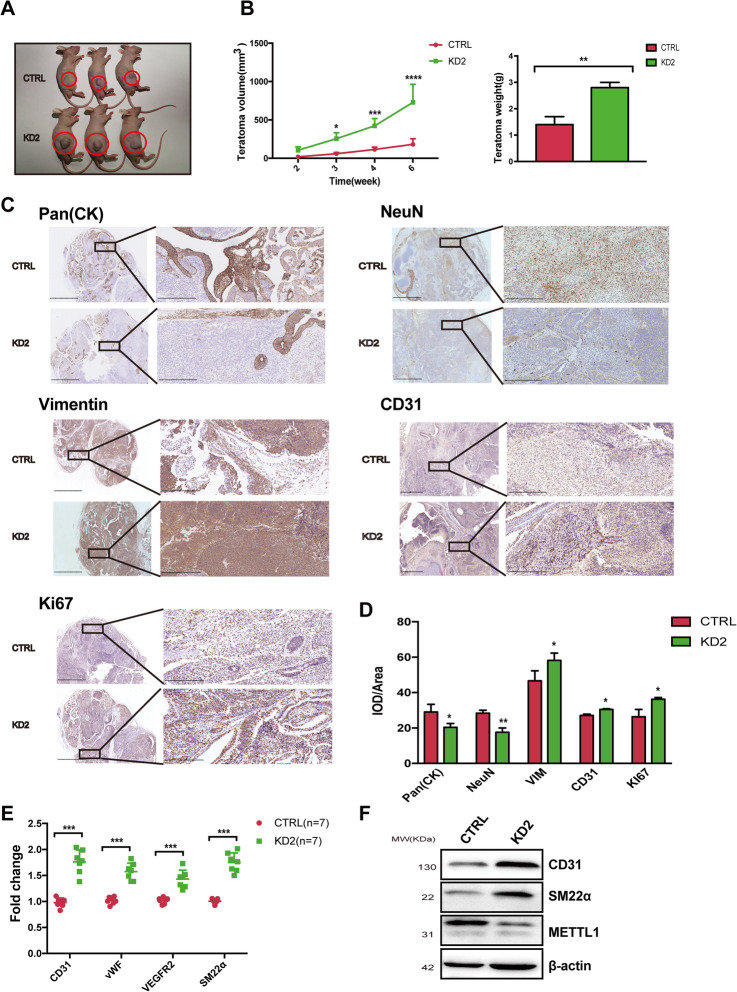
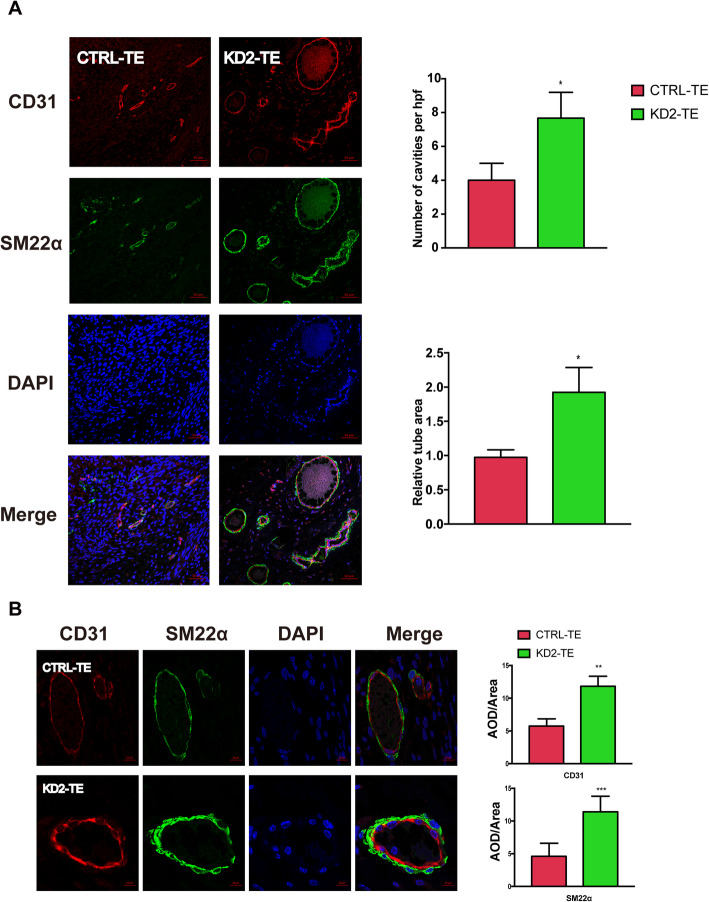
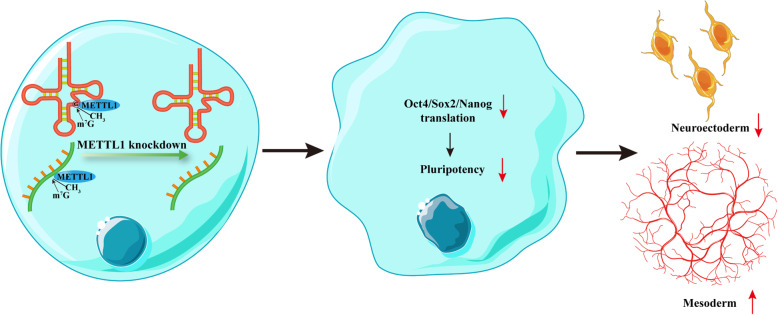
Results: METTL1 silencing resulted in alterations in the global m7G profile in hiPSCs and reduced the translational efficiency of stem cell marker genes. METTL1-KD hiPSCs exhibited reduced pluripotency with slower cell cycling. Moreover, METTL1 silencing accelerates hiPSC differentiation into EBs and promotes the expression of mesoderm-related genes. Similarly, METTL1 knockdown enhances teratoma formation and mesoderm differentiation in vivo by promoting cell proliferation and angiogenesis in nude mice.
Conclusion: Our findings provided novel insight into the critical role of METTL1-mediated m7G modification in the regulation of hiPSC pluripotency and differentiation, as well as its potential roles in vascular development and the treatment of vascular diseases.
Keywords: Differentiation; Human induced pluripotent stem cells (hiPSCs); Mesoderm; N7-methylguanosine (m7G); Pluripotency; Vasculogenesis.
Conflict of interest statement
The authors declare that they have no competing interests
Figures
References
-
- Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131(5):861–872. - PubMed
-
- Yu J, Vodyanik MA, Smuga-Otto K, Antosiewicz-Bourget J, Frane JL, Tian S, Nie J, Jonsdottir GA, Ruotti V, Stewart R, et al. Induced pluripotent stem cell lines derived from human somatic cells. Science. 2007;318(5858):1917–1920. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
