

Type I and Type III Interferons Restrict SARS-CoV-2 Infection of Human Airway Epithelial Cultures
- PMID: 32699094
- PMCID: PMC7495371
- DOI: 10.1128/JVI.00985-20
Type I and Type III Interferons Restrict SARS-CoV-2 Infection of Human Airway Epithelial Cultures
Abstract
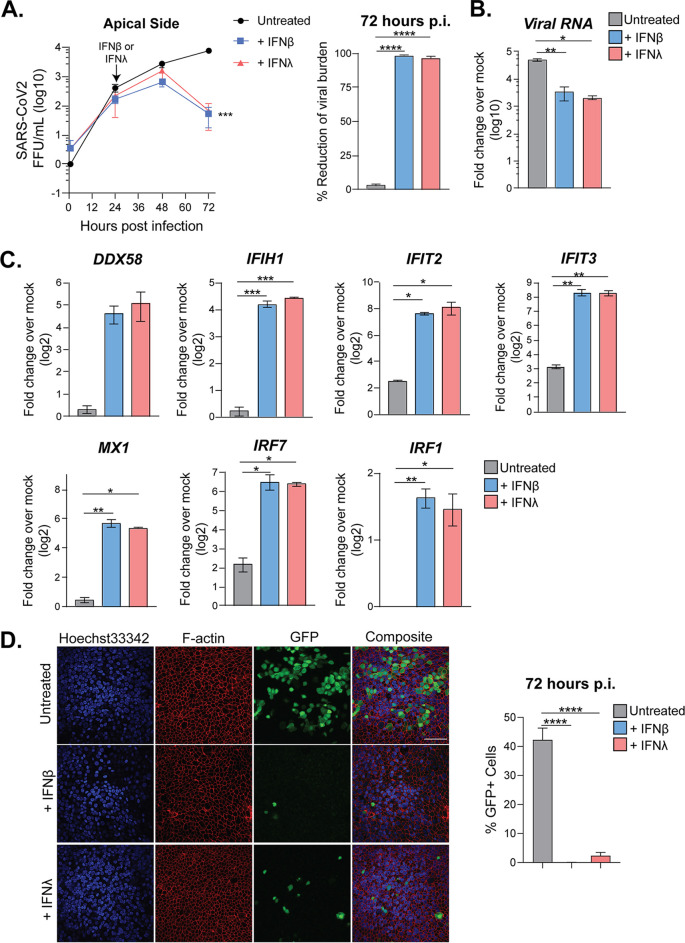
The newly emerged human coronavirus, severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), has caused a pandemic of respiratory illness. Current evidence suggests that severe cases of SARS-CoV-2 are associated with a dysregulated immune response. However, little is known about how the innate immune system responds to SARS-CoV-2. In this study, we modeled SARS-CoV-2 infection using primary human airway epithelial (pHAE) cultures, which are maintained in an air-liquid interface. We found that SARS-CoV-2 infects and replicates in pHAE cultures and is directionally released on the apical, but not basolateral, surface. Transcriptional profiling studies found that infected pHAE cultures had a molecular signature dominated by proinflammatory cytokines and chemokine induction, including interleukin 6 (IL-6), tumor necrosis factor alpha (TNF-α), and CXCL8, and identified NF-κB and ATF-4 as key drivers of this proinflammatory cytokine response. Surprisingly, we observed a complete lack of a type I or III interferon (IFN) response to SARS-CoV-2 infection. However, pretreatment and posttreatment with type I and III IFNs significantly reduced virus replication in pHAE cultures that correlated with upregulation of antiviral effector genes. Combined, our findings demonstrate that SARS-CoV-2 does not trigger an IFN response but is sensitive to the effects of type I and III IFNs. Our studies demonstrate the utility of pHAE cultures to model SARS-CoV-2 infection and that both type I and III IFNs can serve as therapeutic options to treat COVID-19 patients.IMPORTANCE The current pandemic of respiratory illness, COVID-19, is caused by a recently emerged coronavirus named SARS-CoV-2. This virus infects airway and lung cells causing fever, dry cough, and shortness of breath. Severe cases of COVID-19 can result in lung damage, low blood oxygen levels, and even death. As there are currently no vaccines approved for use in humans, studies of the mechanisms of SARS-CoV-2 infection are urgently needed. Our research identifies an excellent system to model SARS-CoV-2 infection of the human airways that can be used to test various treatments. Analysis of infection in this model system found that human airway epithelial cell cultures induce a strong proinflammatory cytokine response yet block the production of type I and III IFNs to SARS-CoV-2. However, treatment of airway cultures with the immune molecules type I or type III interferon (IFN) was able to inhibit SARS-CoV-2 infection. Thus, our model system identified type I or type III IFN as potential antiviral treatments for COVID-19 patients.
Keywords: COVID-19; SARS-CoV-2; cytokines; innate immunity; lung; type I interferon.
Copyright © 2020 American Society for Microbiology.
Figures






References
-
- Zhou P, Yang XL, Wang XG, Hu B, Zhang L, Zhang W, Si HR, Zhu Y, Li B, Huang CL, Chen HD, Chen J, Luo Y, Guo H, Jiang RD, Liu MQ, Chen Y, Shen XR, Wang X, Zheng XS, Zhao K, Chen QJ, Deng F, Liu LL, Yan B, Zhan FX, Wang YY, Xiao GF, Shi ZL. 2020. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579:270–273. doi: 10.1038/s41586-020-2012-7. - DOI - PMC - PubMed
-
- Zhu N, Zhang D, Wang W, Li X, Yang B, Song J, Zhao X, Huang B, Shi W, Lu R, Niu P, Zhan F, Ma X, Wang D, Xu W, Wu G, Gao GF, Tan W, China Novel Coronavirus Investigating and Research Team. 2020. A novel coronavirus from patients with pneumonia in China, 2019. N Engl J Med 382:727–733. doi: 10.1056/NEJMoa2001017. - DOI - PMC - PubMed
-
- World Health Organization. 2020. COVID-19 dashboard. https://covid19.who.int/.
-
- Guan W-J, Ni Z-Y, Hu Y, Liang W-H, Ou C-Q, He J-X, Liu L, Shan H, Lei C-L, Hui DSC, Du B, Li L-J, Zeng G, Yuen K-Y, Chen R-C, Tang C-L, Wang T, Chen P-Y, Xiang J, Li S-Y, Wang J-L, Liang Z-J, Peng Y-X, Wei L, Liu Y, Hu Y-H, Peng P, Wang J-M, Liu J-Y, Chen Z, Li G, Zheng Z-J, Qiu S-Q, Luo J, Ye C-J, Zhu S-Y, Zhong N-S. 2020. Clinical characteristics of coronavirus disease 2019 in China. N Engl J Med 382:1708–1720. doi: 10.1056/NEJMoa2002032. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
