

Pulsed Third-Spin-Assisted Recoupling NMR for Obtaining Long-Range 13C-13C and 15N-13C Distance Restraints
- PMID: 32700540
- PMCID: PMC8324326
- DOI: 10.1021/acs.jpcb.0c04574
Pulsed Third-Spin-Assisted Recoupling NMR for Obtaining Long-Range 13C-13C and 15N-13C Distance Restraints
Abstract
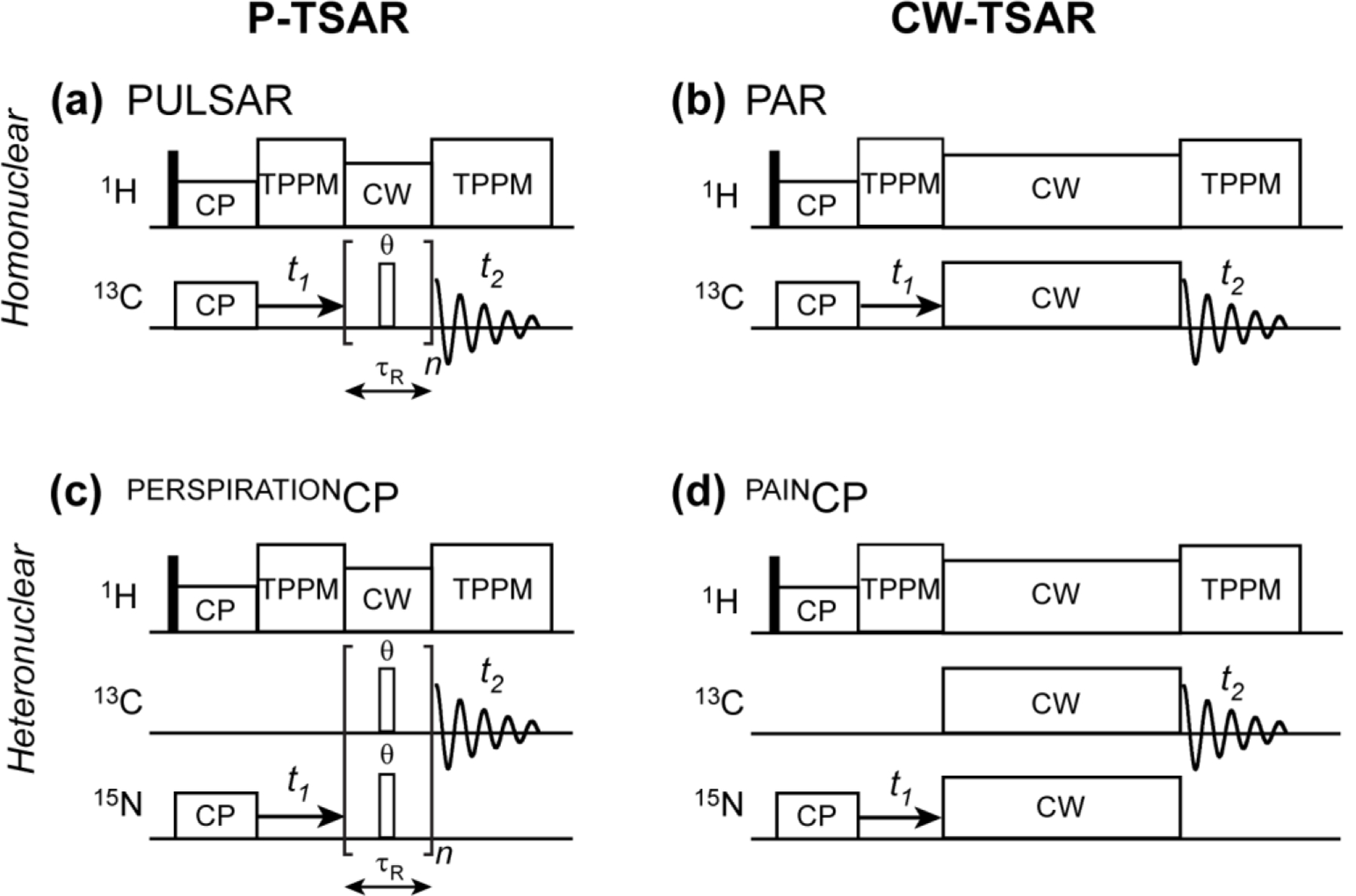
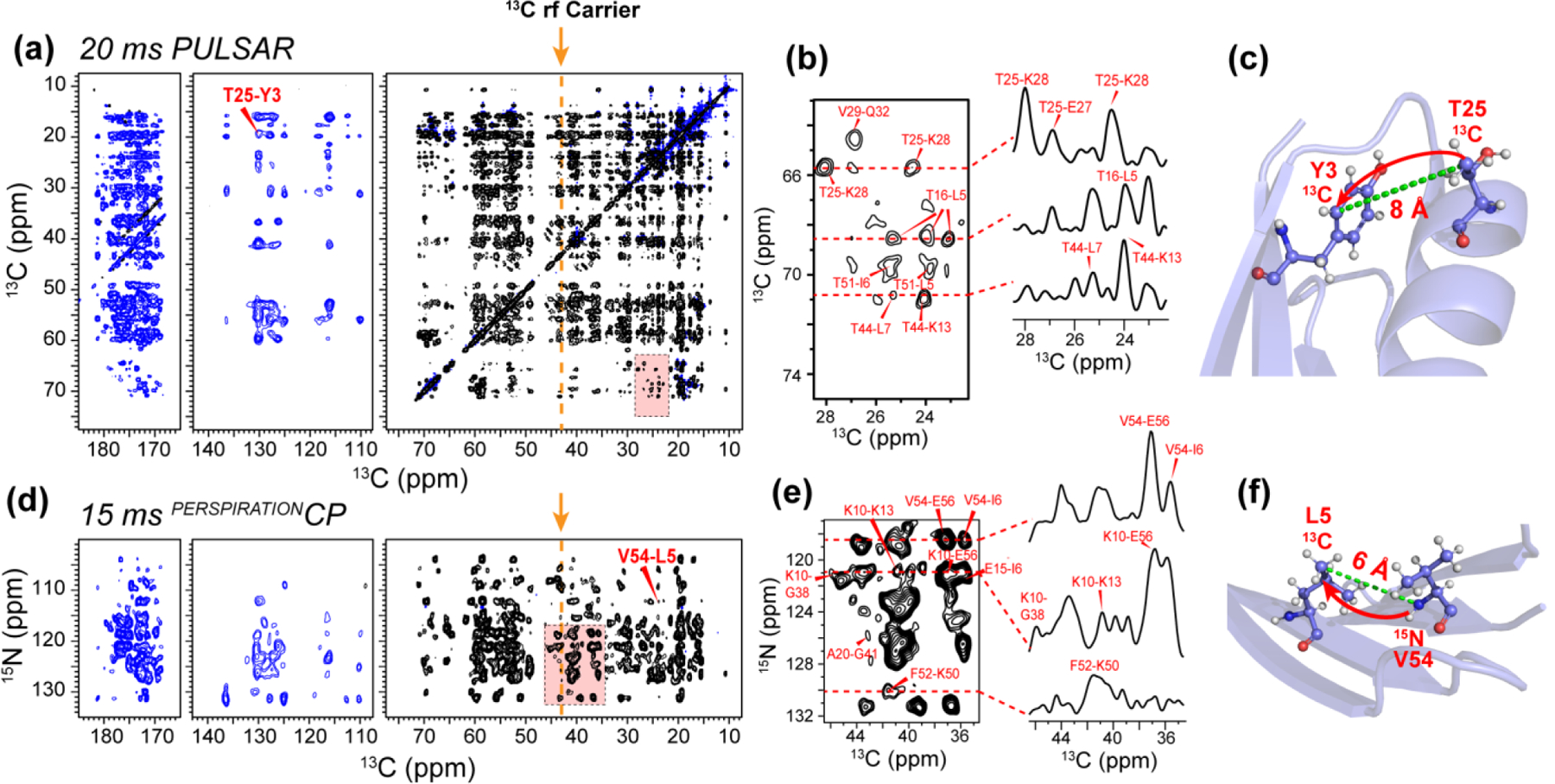
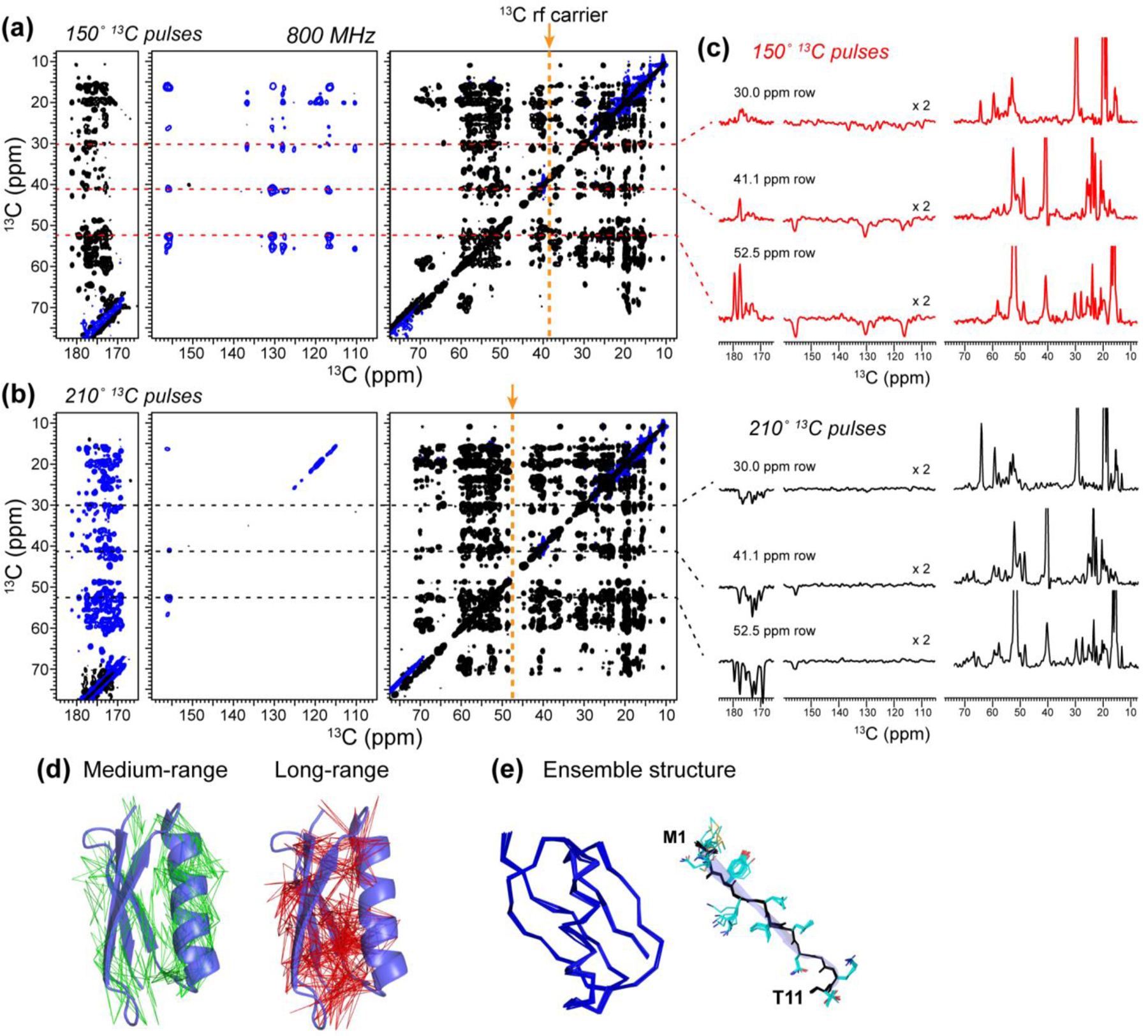
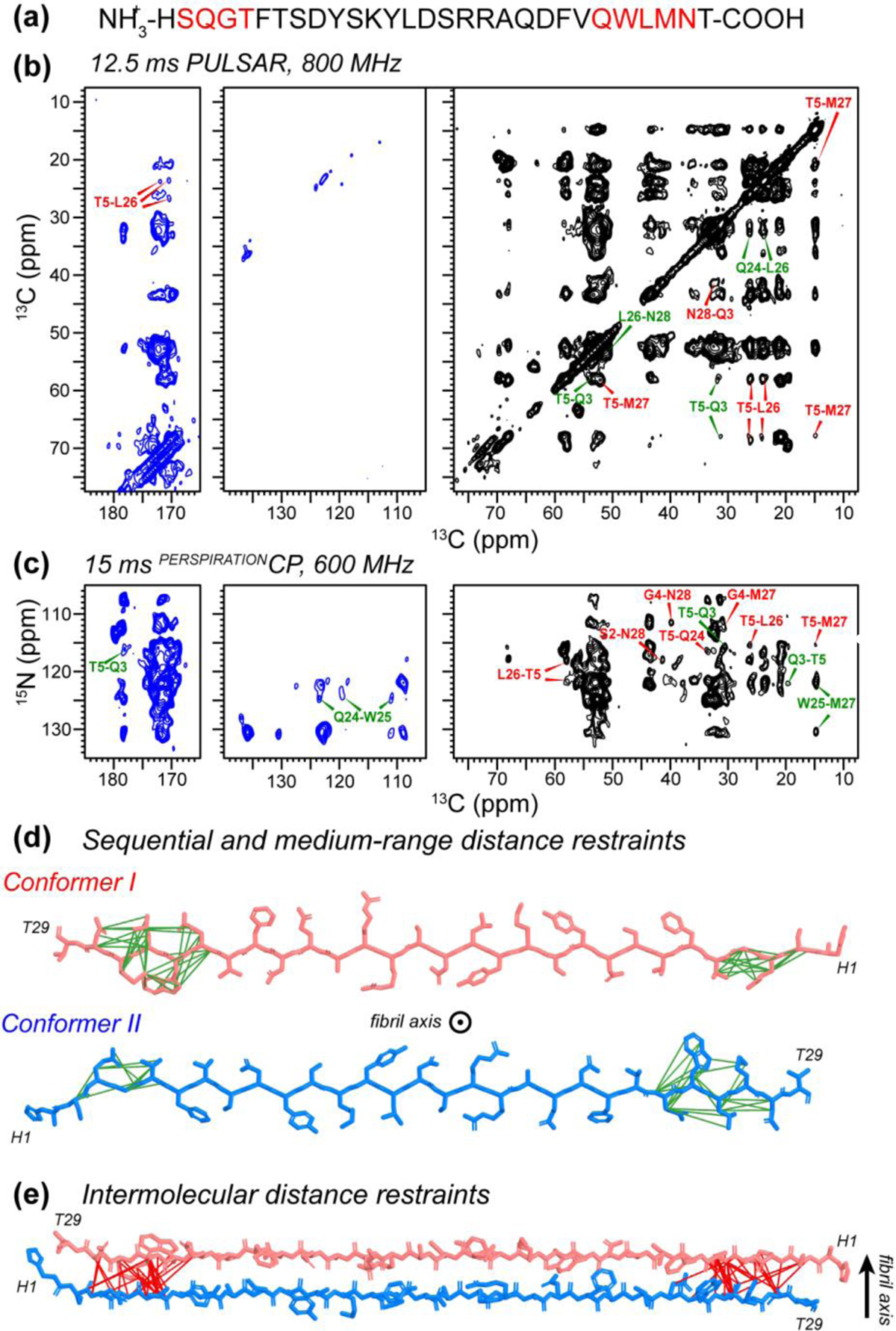
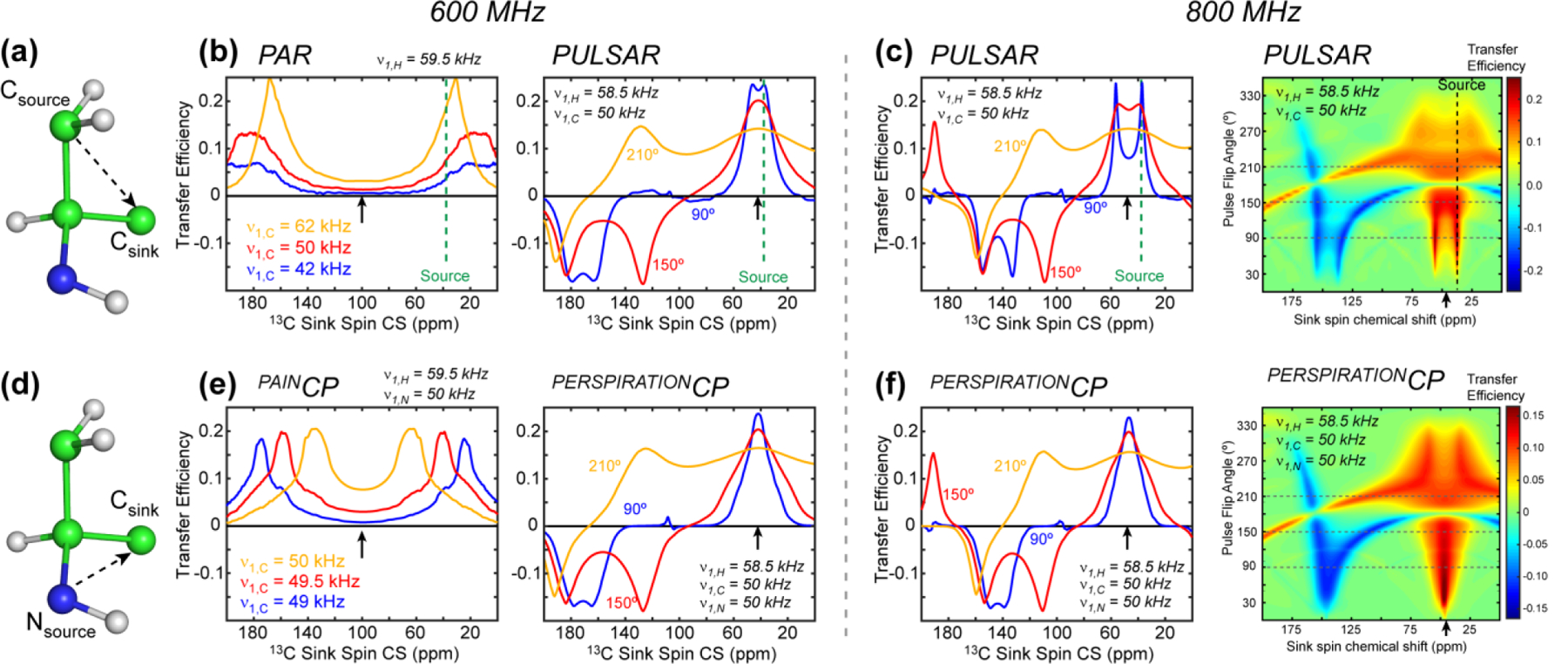
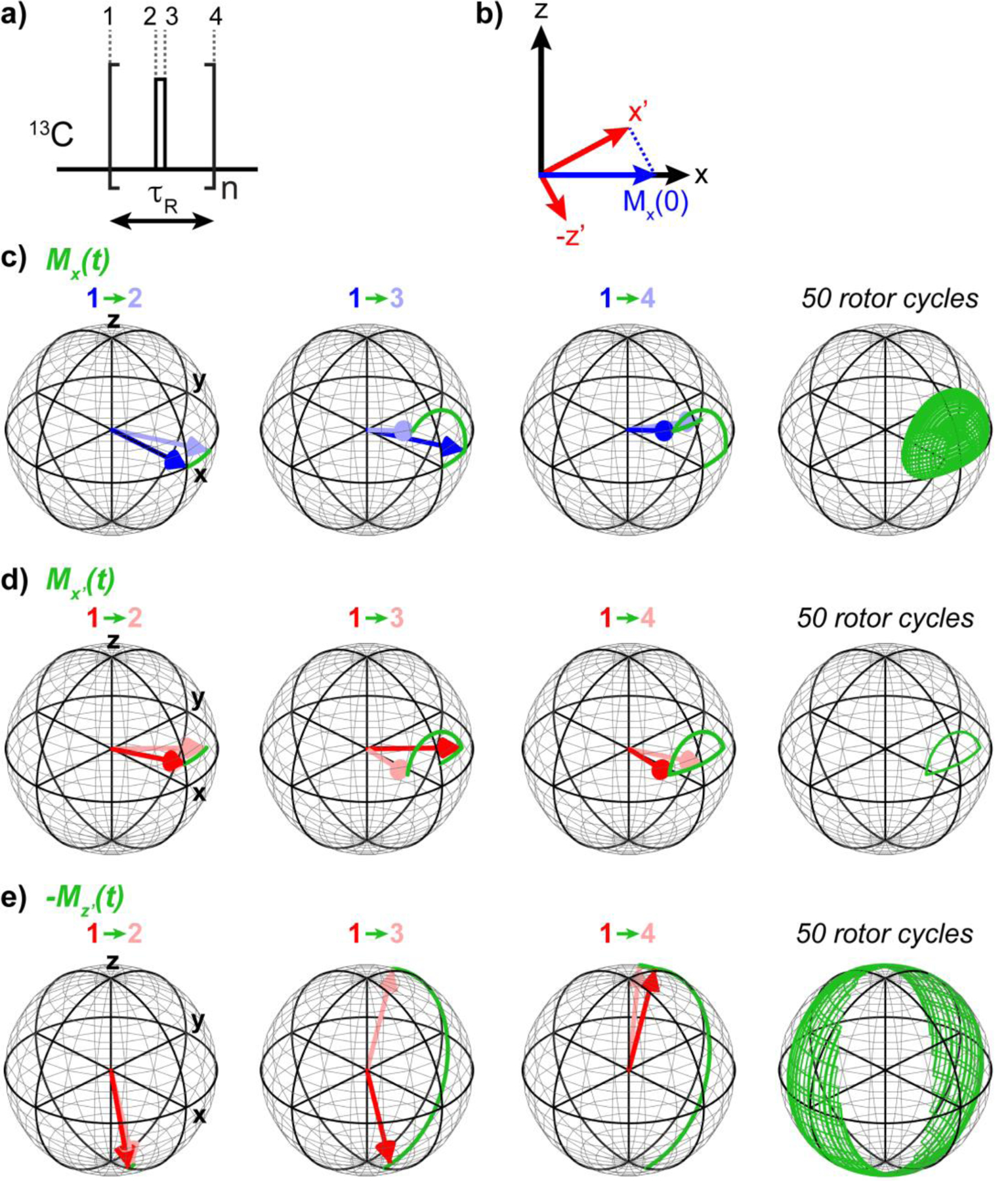
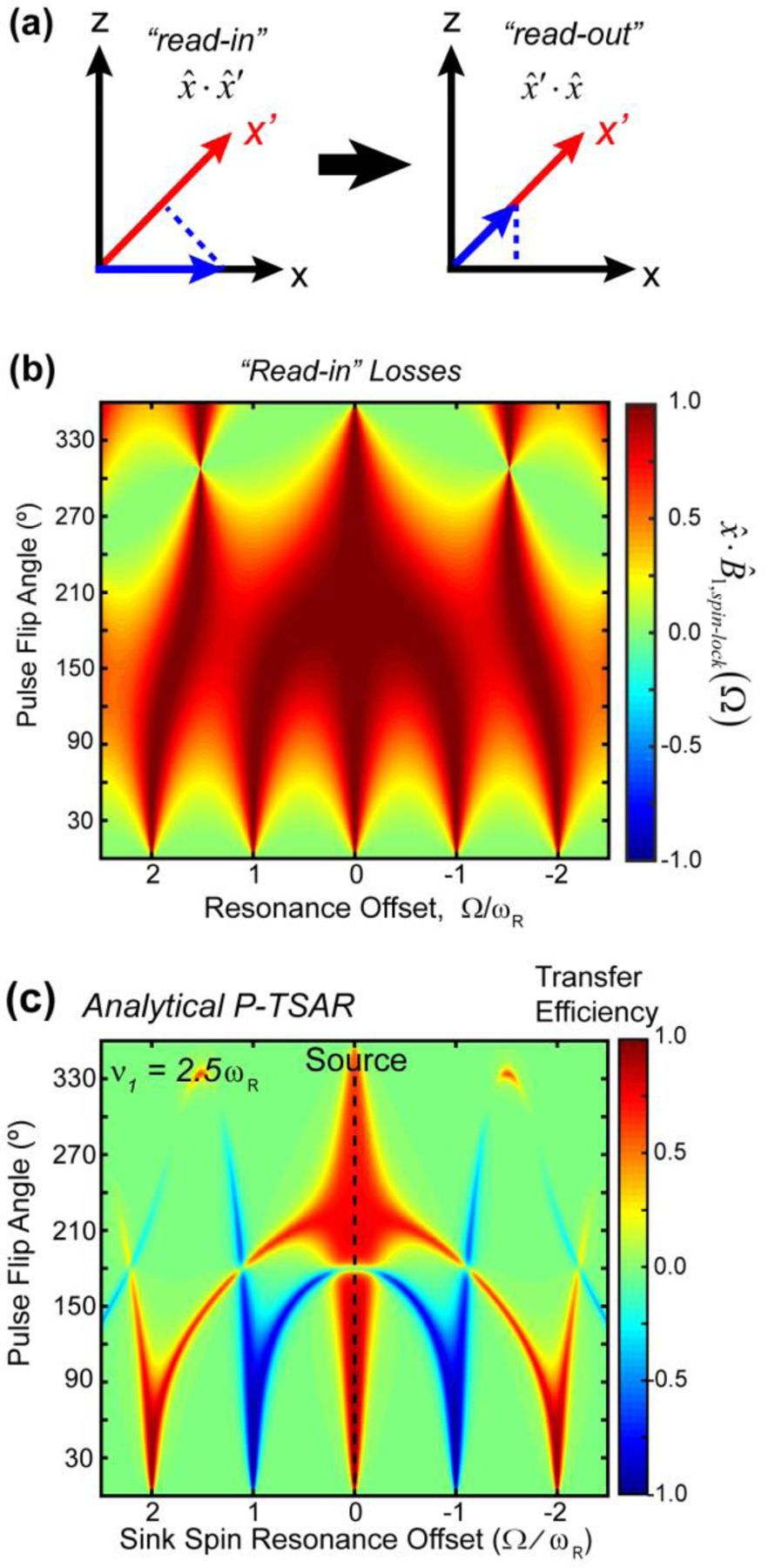
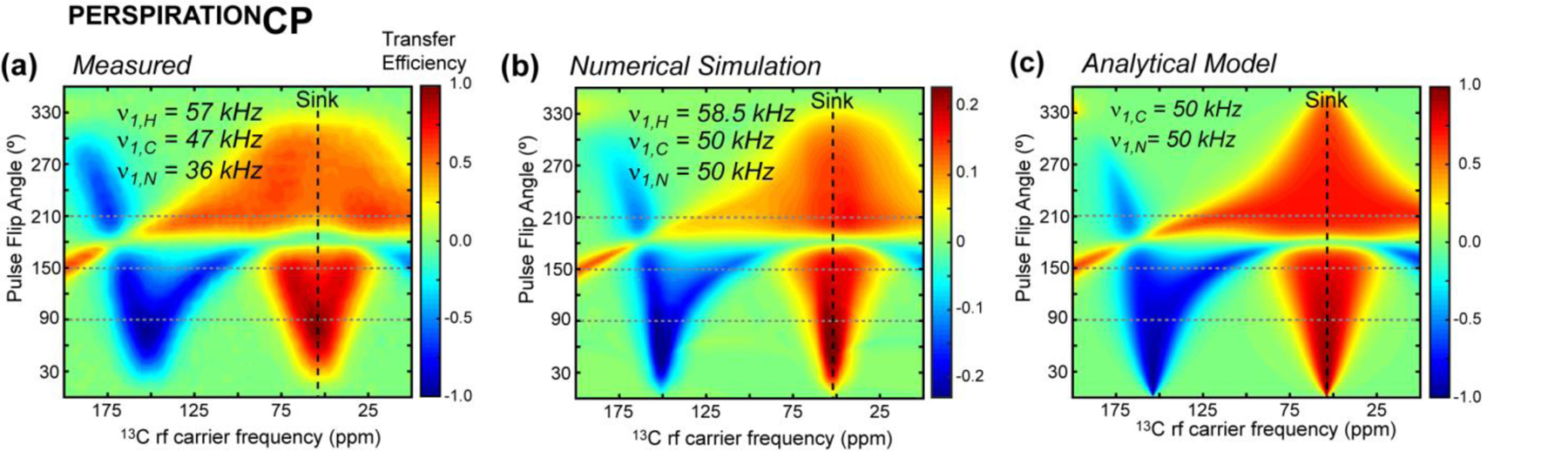
We present a class of pulsed third-spin-assisted recoupling (P-TSAR) magic-angle-spinning solid-state NMR techniques that achieve efficient polarization transfer over long distances to provide important restraints for structure determination. These experiments utilize second-order cross terms between strong 1H-13C and 1H-15N dipolar couplings to achieve 13C-13C and 15N-13C polarization transfer, similar to the principle of continuous-wave (CW) TSAR experiments. However, in contrast to the CW-TSAR experiments, these P-TSAR experiments require much less radiofrequency (rf) energy and allow a much simpler routine for optimizing the rf field strength. We call the technique PULSAR (pulsed proton-assisted recoupling) for homonuclear spin pairs. For heteronuclear spin pairs, we improve the recently introduced PERSPIRATIONCP (proton-enhanced rotor-echo short pulse irradiation cross-polarization) experiment by shifting the pulse positions and removing the z-filters, which significantly broaden the bandwidth and increase the efficiency of polarization transfer. We demonstrate the PULSAR and PERSPIRATIONCP techniques on the model protein GB1 and found cross peaks for distances as long as 10 and 8 Å for 13C-13C and 15N-13C spin pairs, respectively. We then apply these methods to the amyloid fibrils formed by the peptide hormone glucagon and show that long-range correlation peaks are readily observed to constrain intermolecular packing in this cross-β fibril. We provide an analytical model for the PULSAR and PERSPIRATIONCP experiments to explain the measured and simulated chemical shift dependence and pulse flip angle dependence of polarization transfer. These two techniques are useful for measuring long-range distance restraints to determine the three-dimensional structures of proteins and other biological macromolecules.
Figures
References
-
- Wang SL; Munro RA; Shi LC; Kawamura I; Okitsu T; Wada A; Kim SY; Jung KH; Brown LS; Ladizhansky V., Solid-state NMR spectroscopy structure determination of a lipid-embedded heptahelical membrane protein. Nature Methods 2013, 10, 1007–1012. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
