

Sensitivity of allyl isothiocyanate to induce apoptosis via ER stress and the mitochondrial pathway upon ROS production in colorectal adenocarcinoma cells
- PMID: 32700751
- PMCID: PMC7448487
- DOI: 10.3892/or.2020.7700
Sensitivity of allyl isothiocyanate to induce apoptosis via ER stress and the mitochondrial pathway upon ROS production in colorectal adenocarcinoma cells
Abstract
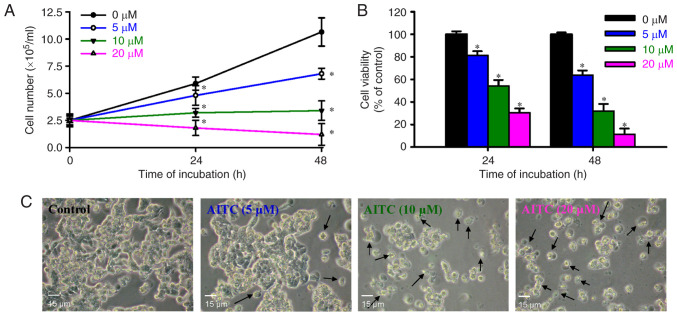
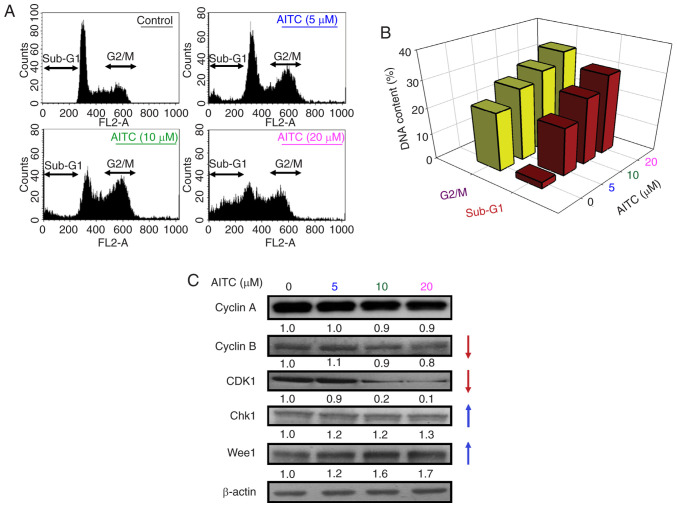
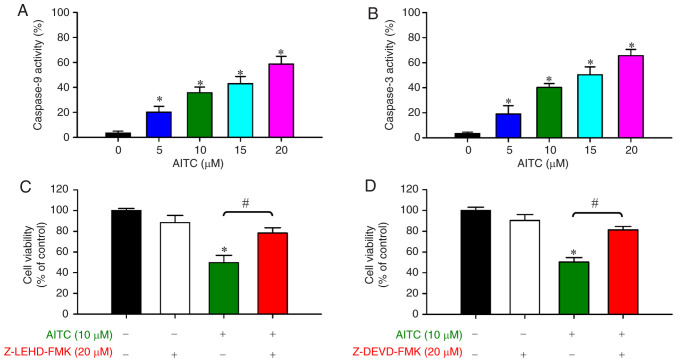
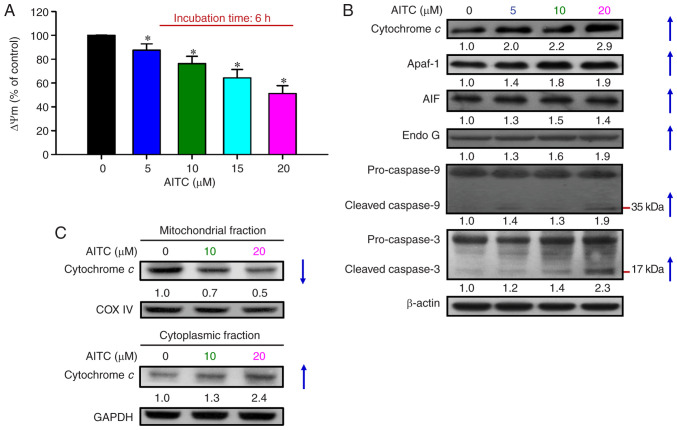
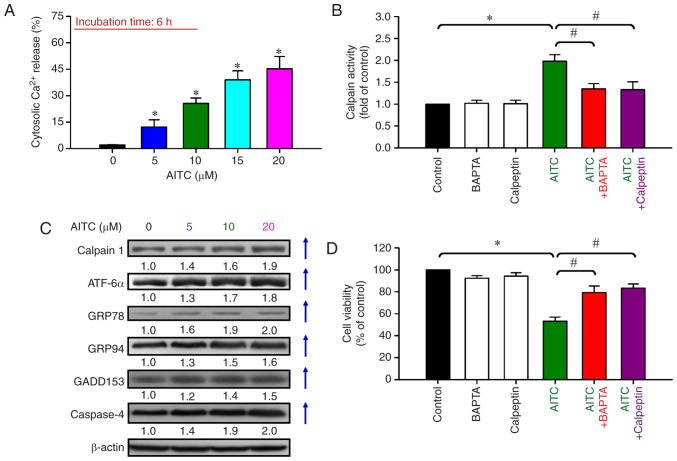
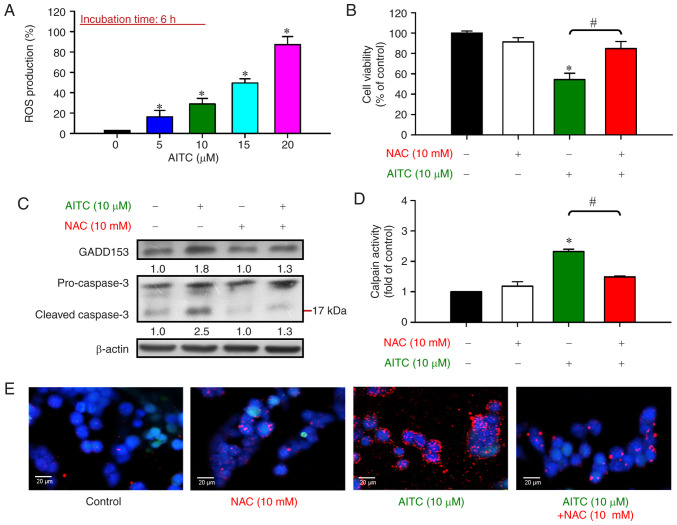
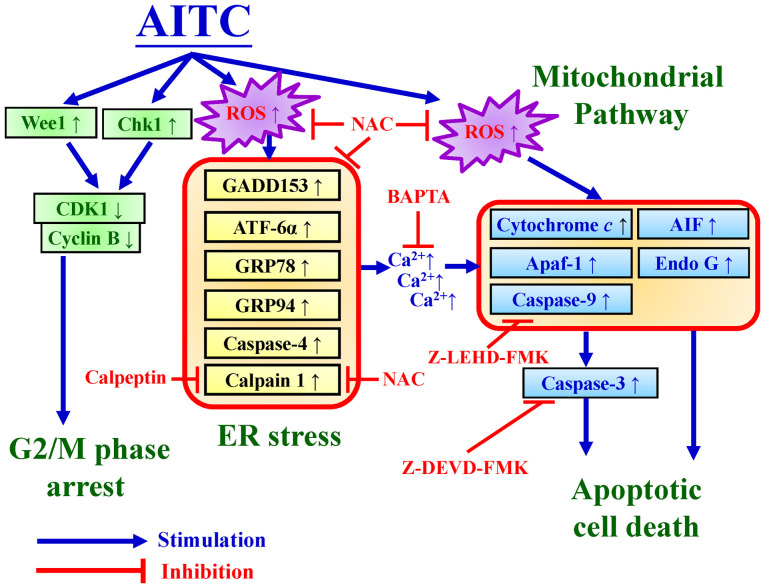
Allyl isothiocyanate (AITC), a bioactive phytochemical compound that is a constituent of dietary cruciferous vegetables, possesses promising chemopreventive and anticancer effects. However, reports of AITC exerting antitumor effects on apoptosis induction of colorectal cancer (CRC) cells in vitro are not well elucidated. The present study focused on the functional mechanism of the endoplasmic reticulum (ER) stress‑based apoptotic machinery induced by AITC in human colorectal cancer HT‑29 cells. Our results indicated that AITC decreased cell growth and number, reduced viability, and facilitated morphological changes of apoptotic cell death. DNA analysis by flow cytometry showed G2/M phase arrest, and alterations in the modulated protein levels caused by AITC were detected via western blot analysis. AITC also triggered vital intrinsic apoptotic factors (caspase‑9/caspase‑3 activity), disrupted mitochondrial membrane potential, and stimulated mitochondrial‑related apoptotic molecules (e.g., cytochrome c, apoptotic protease activating factor 1, apoptosis‑inducing factor, and endonuclease G). Additionally, AITC prompted induced cytosolic Ca2+ release and Ca2+‑dependent ER stress‑related signals, such as calpain 1, activating transcription factor 6α, glucose‑regulated proteins 78 and 94, growth arrest‑ and DNA damage‑inducible protein 153 (GADD153), and caspase‑4. The level of reactive oxygen species (ROS) production was found to induce the hallmark of ER stress GADD153, proapoptotic marker caspase‑3, and calpain activity after AITC treatment. Our findings showed for the first time that AITC induced G2/M phase arrest and apoptotic death via ROS‑based ER stress and the intrinsic pathway (mitochondrial‑dependent) in HT‑29 cells. Overall, AITC may exert an epigenetic effect and is a potential bioactive compound for CRC treatment.
Keywords: allyl isothiocyanate; colon cancer cells; apoptosis; ER stress; mitochondria; ROS production.
Figures
References
-
- Afrin S, Giampieri F, Gasparrini M, Forbes-Hernández TY, Cianciosi D, Reboredo-Rodriguez P, Zhang J, Manna PP, Daglia M, Atanasov AG, Battino M. Dietary phytochemicals in colorectal cancer prevention and treatment: A focus on the molecular mechanisms involved. Biotechnol Adv. 2020;38:107322. doi: 10.1016/j.biotechadv.2018.11.011. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
