

Circadian Photoentrainment in Mice and Humans
- PMID: 32708259
- PMCID: PMC7408241
- DOI: 10.3390/biology9070180
Circadian Photoentrainment in Mice and Humans
Abstract
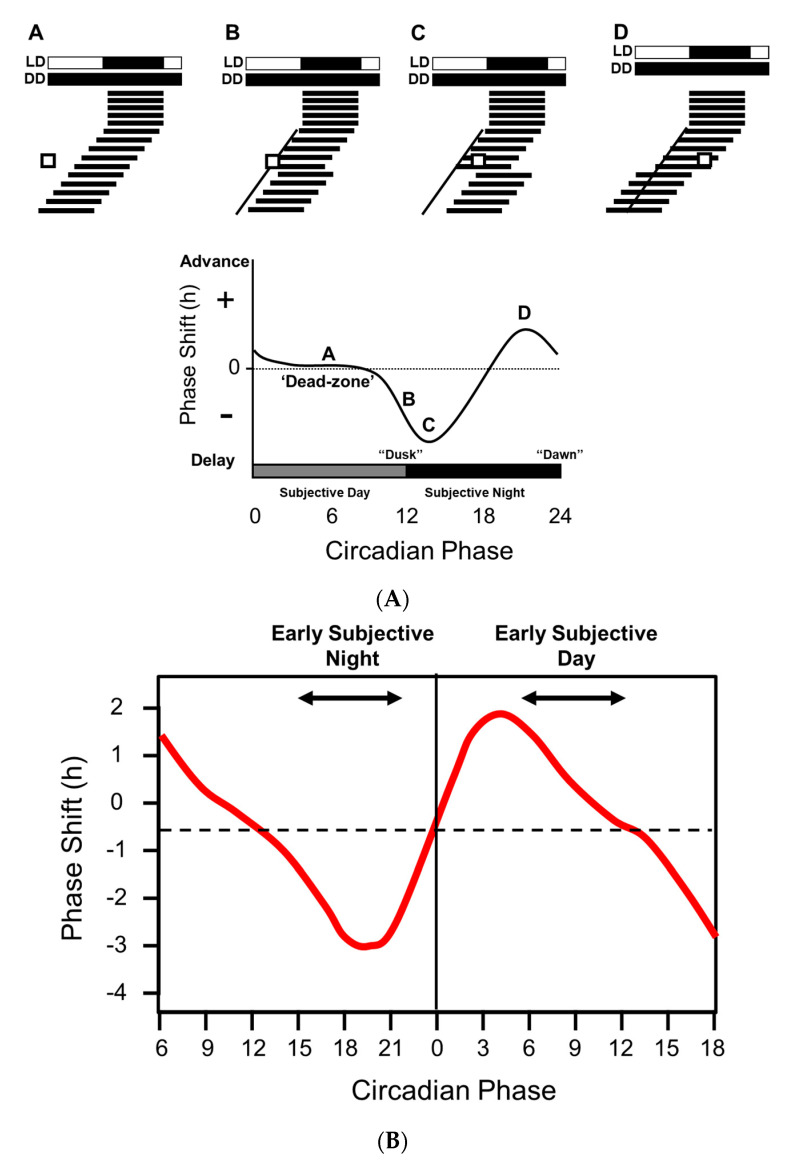
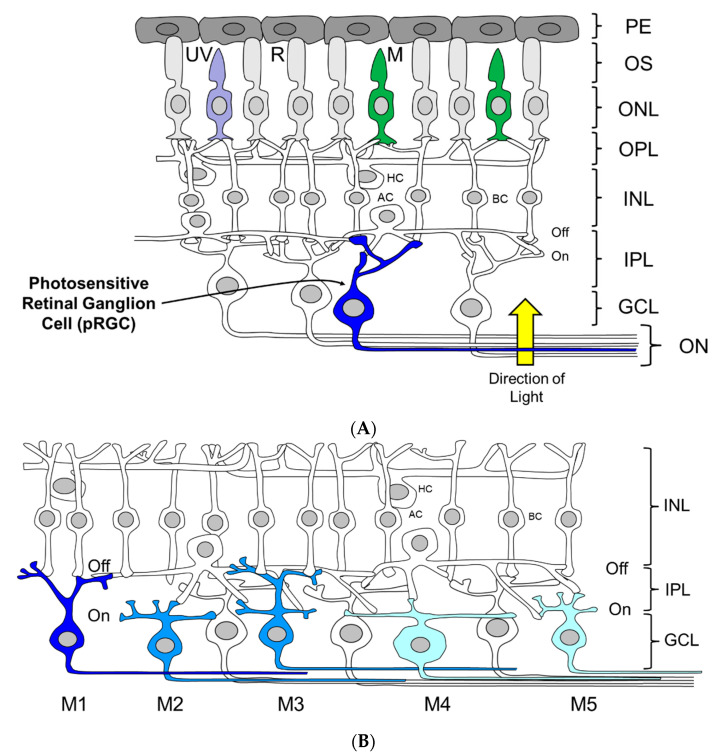
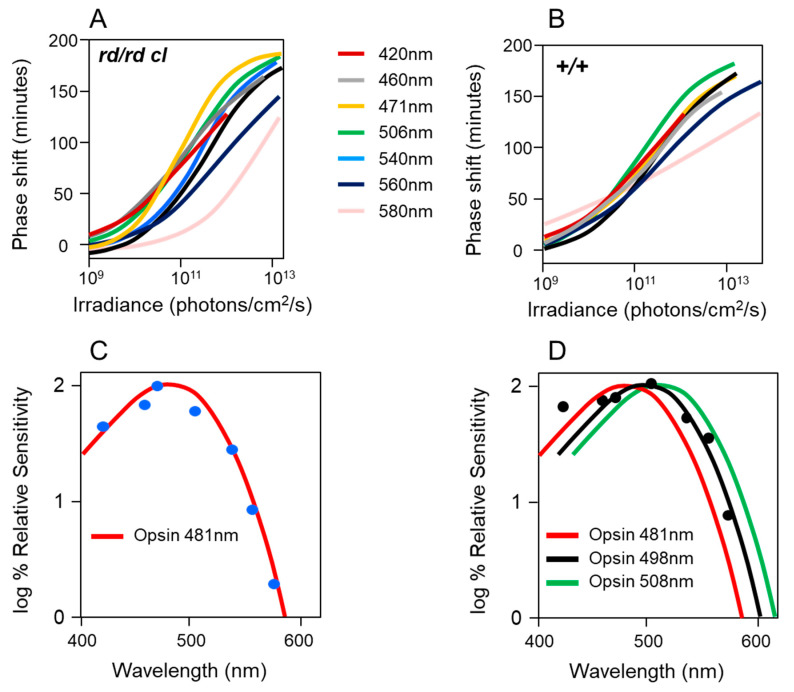
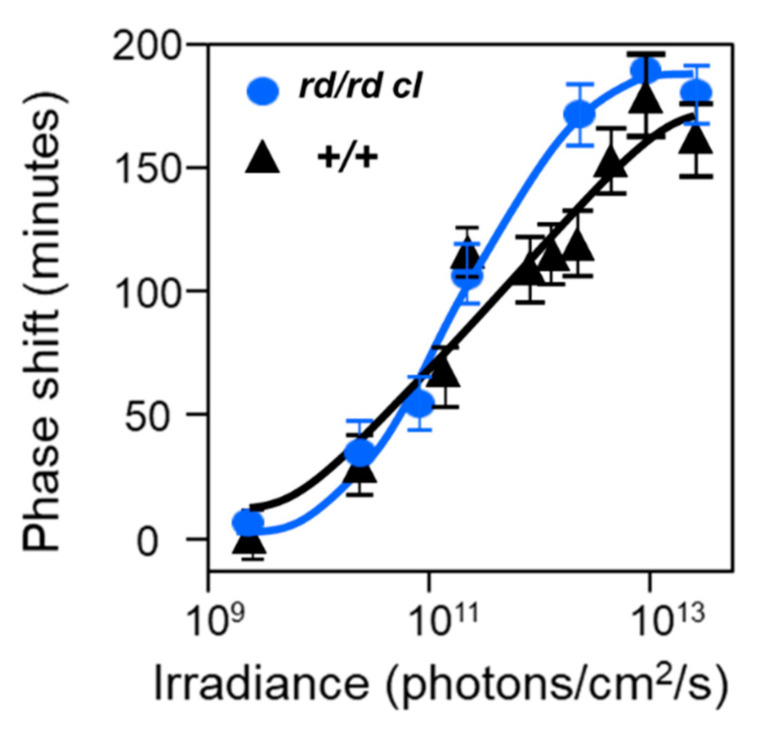
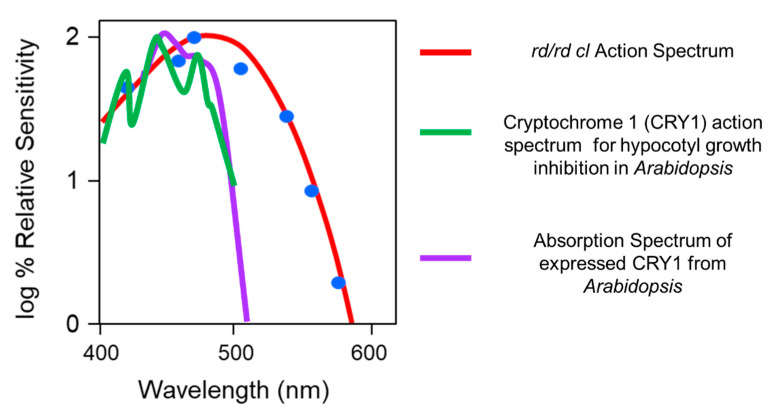
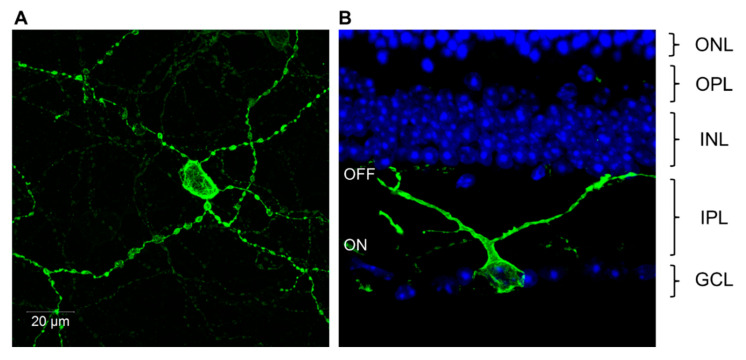
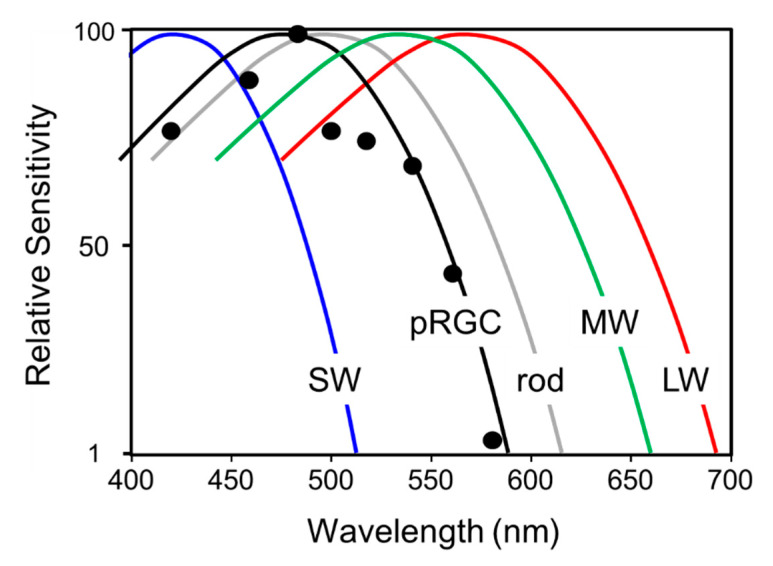
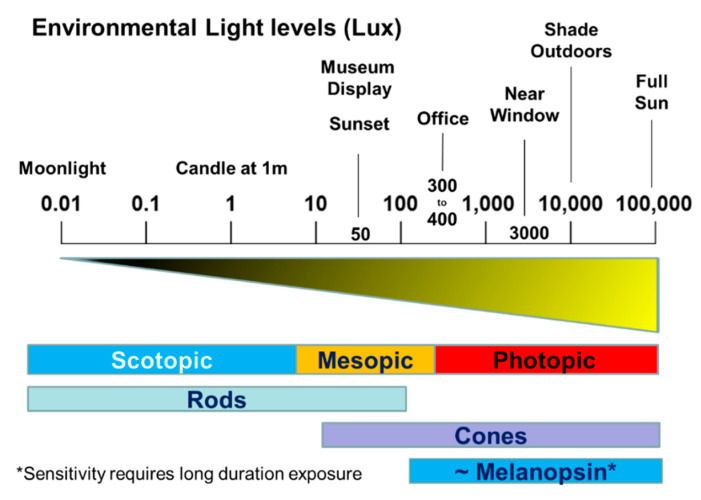
Light around twilight provides the primary entrainment signal for circadian rhythms. Here we review the mechanisms and responses of the mouse and human circadian systems to light. Both utilize a network of photosensitive retinal ganglion cells (pRGCs) expressing the photopigment melanopsin (OPN4). In both species action spectra and functional expression of OPN4 in vitro show that melanopsin has a λmax close to 480 nm. Anatomical findings demonstrate that there are multiple pRGC sub-types, with some evidence in mice, but little in humans, regarding their roles in regulating physiology and behavior. Studies in mice, non-human primates and humans, show that rods and cones project to and can modulate the light responses of pRGCs. Such an integration of signals enables the rods to detect dim light, the cones to detect higher light intensities and the integration of intermittent light exposure, whilst melanopsin measures bright light over extended periods of time. Although photoreceptor mechanisms are similar, sensitivity thresholds differ markedly between mice and humans. Mice can entrain to light at approximately 1 lux for a few minutes, whilst humans require light at high irradiance (>100's lux) and of a long duration (>30 min). The basis for this difference remains unclear. As our retinal light exposure is highly dynamic, and because photoreceptor interactions are complex and difficult to model, attempts to develop evidence-based lighting to enhance human circadian entrainment are very challenging. A way forward will be to define human circadian responses to artificial and natural light in the "real world" where light intensity, duration, spectral quality, time of day, light history and age can each be assessed.
Keywords: circadian; entrainment; human; melanopsin (OPN4); mouse; photoreceptor.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Daan S., Pittendrigh C.S. A functional analysis of circadian pacemakers in nocturnal rodents. II. The variabilityof phase response curves. J. Comp. Physiol. 1976;106:253–266. doi: 10.1007/BF01417857. - DOI
Publication types
LinkOut - more resources
Full Text Sources
Research Materials
