

Epigenetics as an Evolutionary Tool for Centromere Flexibility
- PMID: 32708654
- PMCID: PMC7397245
- DOI: 10.3390/genes11070809
Epigenetics as an Evolutionary Tool for Centromere Flexibility
Abstract
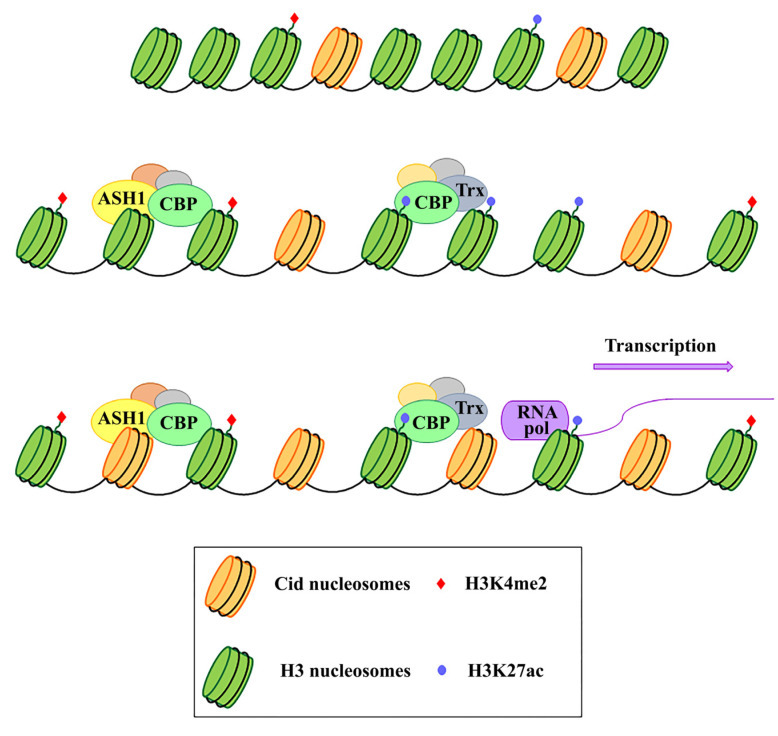
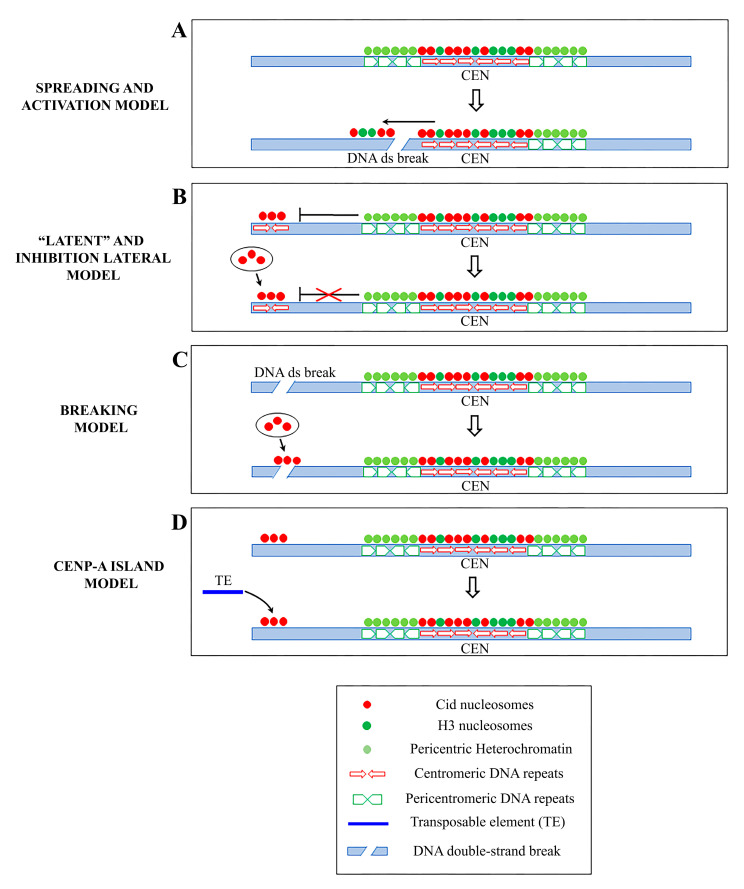
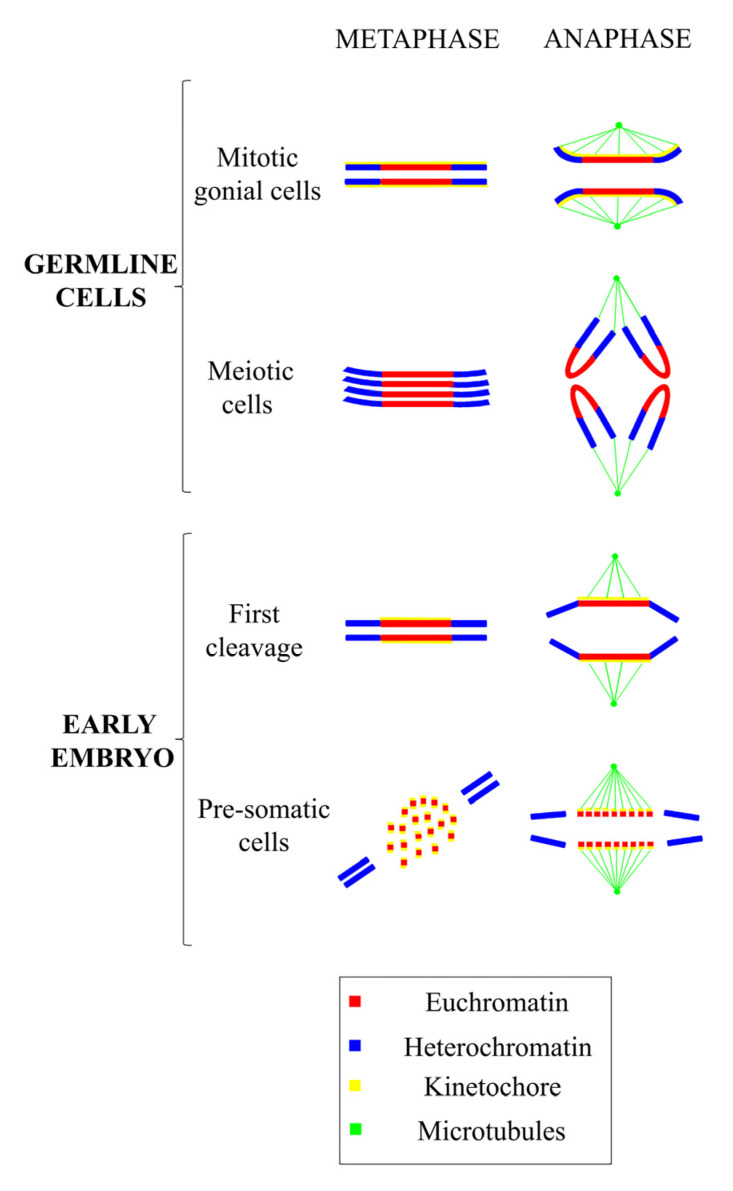
Centromeres are the complex structures responsible for the proper segregation of chromosomes during cell division. Structural or functional alterations of the centromere cause aneuploidies and other chromosomal aberrations that can induce cell death with consequences on health and survival of the organism as a whole. Because of their essential function in the cell, centromeres have evolved high flexibility and mechanisms of tolerance to preserve their function following stress, whether it is originating from within or outside the cell. Here, we review the main epigenetic mechanisms of centromeres' adaptability to preserve their functional stability, with particular reference to neocentromeres and holocentromeres. The centromere position can shift in response to altered chromosome structures, but how and why neocentromeres appear in a given chromosome region are still open questions. Models of neocentromere formation developed during the last few years will be hereby discussed. Moreover, we will discuss the evolutionary significance of diffuse centromeres (holocentromeres) in organisms such as nematodes. Despite the differences in DNA sequences, protein composition and centromere size, all of these diverse centromere structures promote efficient chromosome segregation, balancing genome stability and adaptability, and ensuring faithful genome inheritance at each cellular generation.
Keywords: CENP-A; centromere; centromere evolution; holocentromere; neocentromere; repetitive sequences.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
