

Structural Basis of Human Helicase DDX21 in RNA Binding, Unwinding, and Antiviral Signal Activation
- PMID: 32714761
- PMCID: PMC7375243
- DOI: 10.1002/advs.202000532
Structural Basis of Human Helicase DDX21 in RNA Binding, Unwinding, and Antiviral Signal Activation
Abstract
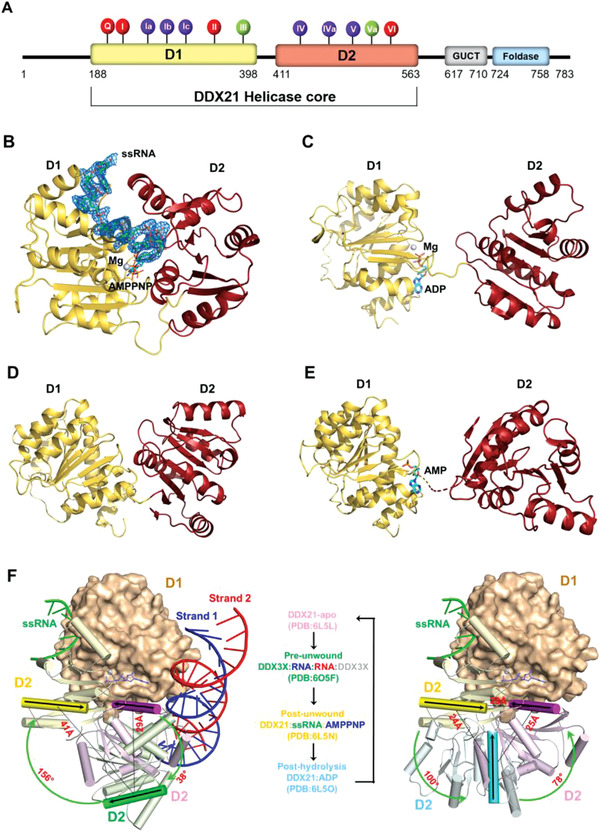
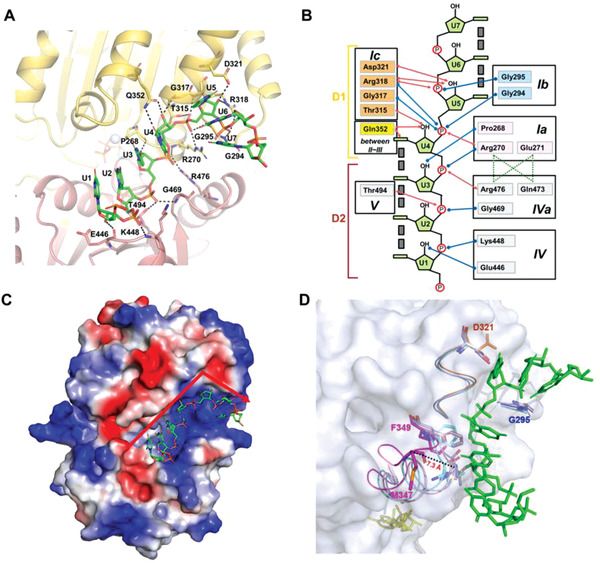
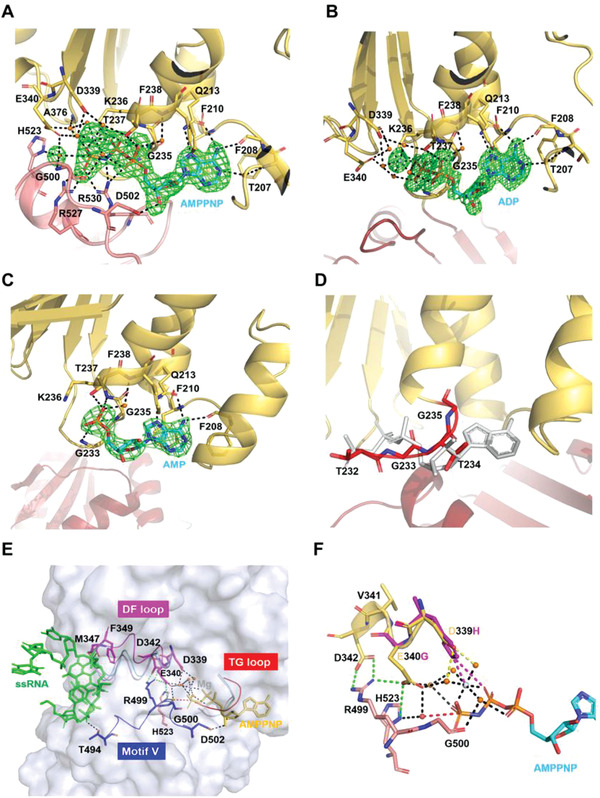
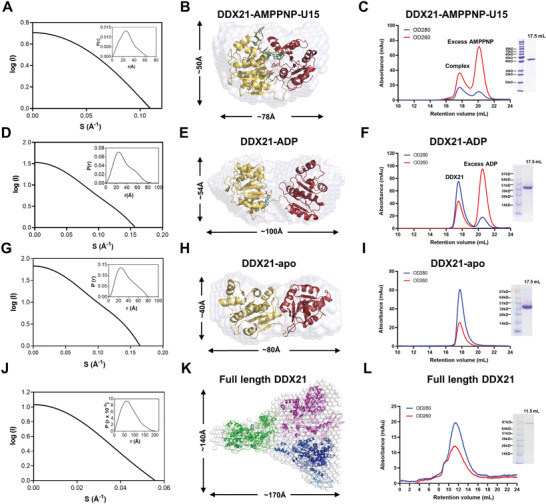
RNA helicase DDX21 plays vital roles in ribosomal RNA biogenesis, transcription, and the regulation of host innate immunity during virus infection. How DDX21 recognizes and unwinds RNA and how DDX21 interacts with virus remain poorly understood. Here, crystal structures of human DDX21 determined in three distinct states are reported, including the apo-state, the AMPPNP plus single-stranded RNA (ssRNA) bound pre-hydrolysis state, and the ADP-bound post-hydrolysis state, revealing an open to closed conformational change upon RNA binding and unwinding. The core of the RNA unwinding machinery of DDX21 includes one wedge helix, one sensor motif V and the DEVD box, which links the binding pockets of ATP and ssRNA. The mutant D339H/E340G dramatically increases RNA binding activity. Moreover, Hill coefficient analysis reveals that DDX21 unwinds double-stranded RNA (dsRNA) in a cooperative manner. Besides, the nonstructural (NS1) protein of influenza A inhibits the ATPase and unwinding activity of DDX21 via small RNAs, which cooperatively assemble with DDX21 and NS1. The structures illustrate the dynamic process of ATP hydrolysis and RNA unwinding for RNA helicases, and the RNA modulated interaction between NS1 and DDX21 generates a fresh perspective toward the virus-host interface. It would benefit in developing therapeutics to combat the influenza virus infection.
Keywords: ATPases; DDX21; RNA helicases; crystal structures; viral protein NS1.
© 2020 The Authors. Published by WILEY‐VCH Verlag GmbH & Co. KGaA, Weinheim.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
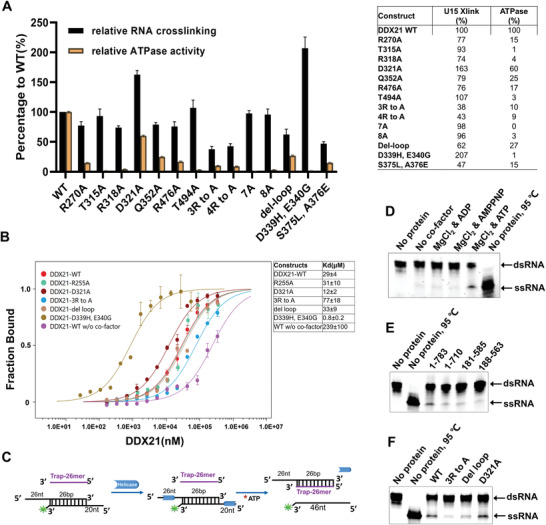
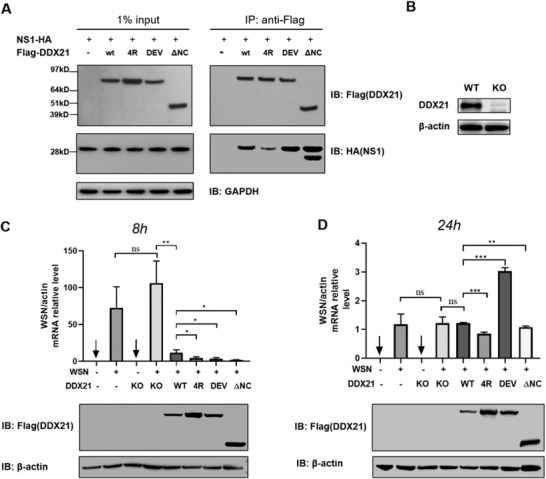
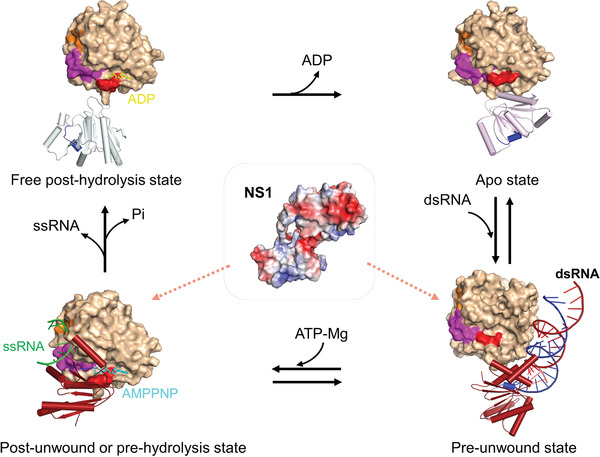
References
-
- a) Linder P., Nucleic Acids Res. 2006, 34, 4168; - PMC - PubMed
- b) Martin R., Straub A. U., Doebele C., Bohnsack M. T., RNA Biol. 2013, 10, 4; - PMC - PubMed
- c) Sloan K. E., Leisegang M. S., Doebele C., Ram A. S., Simm S., Safferthal C., Kretschmer J., Schorge T., Markoutsa S., Haag S., Karas M., Ebersberger I., Schleiff E., Watkins N. J., Bohnsack M. T., Nucleic Acids Res. 2015, 43, 553. - PMC - PubMed
-
- Fuller‐pace F. V, Nicol S. M., Methods Enzymol. 2012, 511, 347. - PubMed
-
- Steimer L., Klostermeier D., RNA Biol. 2012, 9, 751. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous