

Glutaminase Inhibition on NSCLC Depends on Extracellular Alanine Exploitation
- PMID: 32718002
- PMCID: PMC7465377
- DOI: 10.3390/cells9081766
Glutaminase Inhibition on NSCLC Depends on Extracellular Alanine Exploitation
Abstract
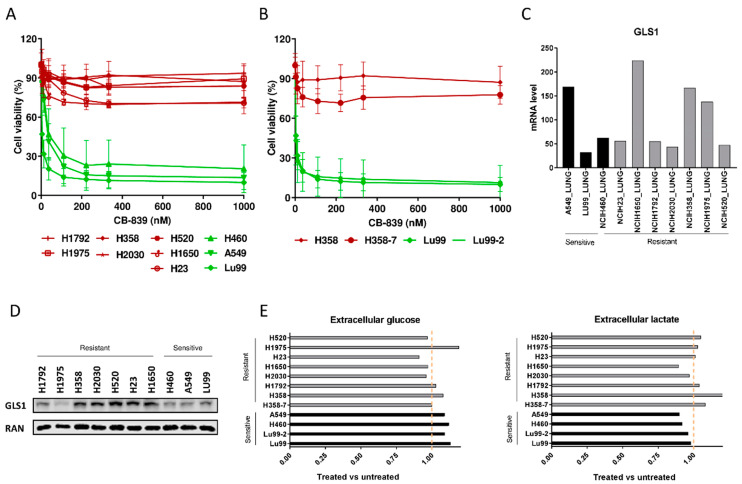
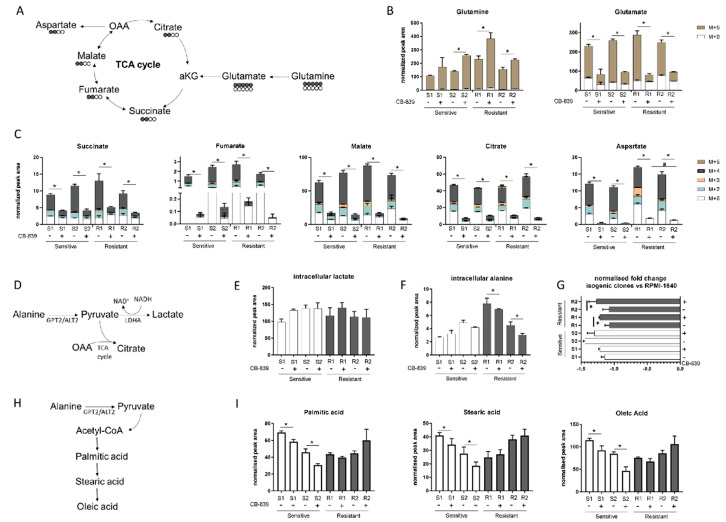
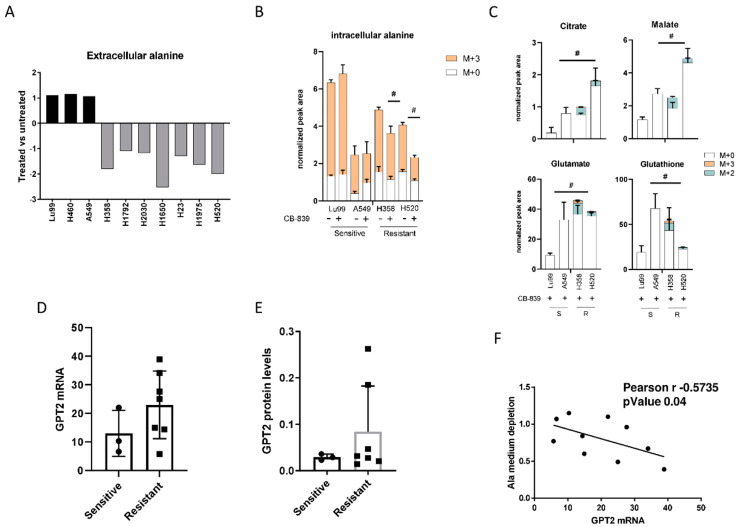
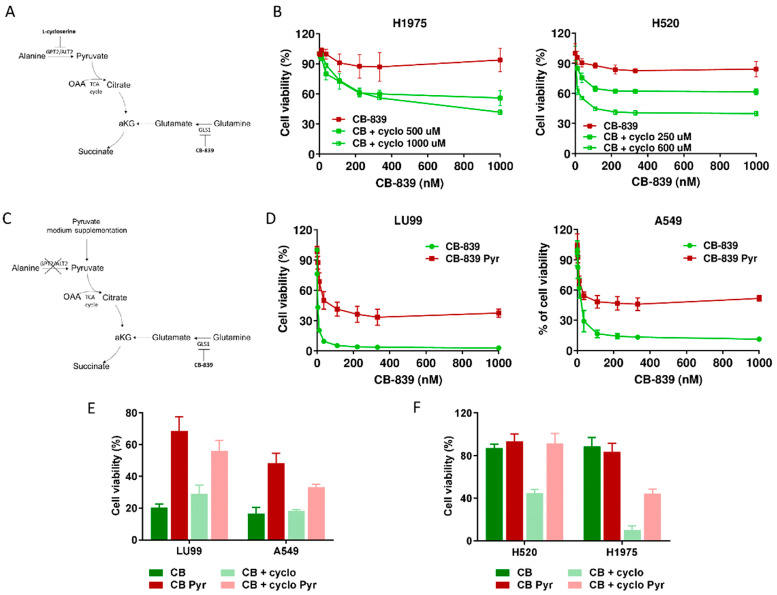
Non-small-cell lung cancer (NSCLC) cell lines vary in their sensitivity to glutaminase inhibitors, so it is important to identify the metabolic assets underling their efficacy in cancer cells. Even though specific genetic lesions such as in KRAS and LKB1 have been associated with reliance on glutamine for their metabolic needs, we found no distinction between glutaminase inhibitor CB-839 sensitivity and resistant phenotypes in NSCLC cells with or without these genetic alterations. We demonstrated the close relationship between environmental alanine uptake and catabolism. This response depended on the individual cell's ability to employ alanine aminotransferase (GPT2) to compensate the reduced glutamate availability. It may, therefore, be useful to determine GPT2 levels to predict which NSCLC patients would benefit most from glutaminase inhibitor treatment.
Keywords: NSCLC; alanine aminotransferase; alanine uptake; cancer cell resistance; glutaminase inhibition; metabolism.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
