

Elevated energy requirement of cone photoreceptors
- PMID: 32719136
- PMCID: PMC7431031
- DOI: 10.1073/pnas.2001776117
Elevated energy requirement of cone photoreceptors
Abstract
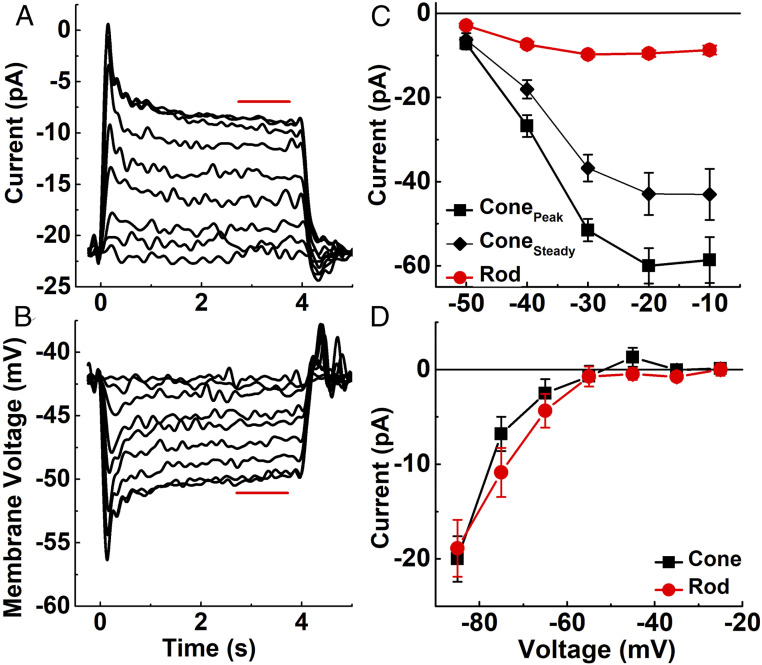
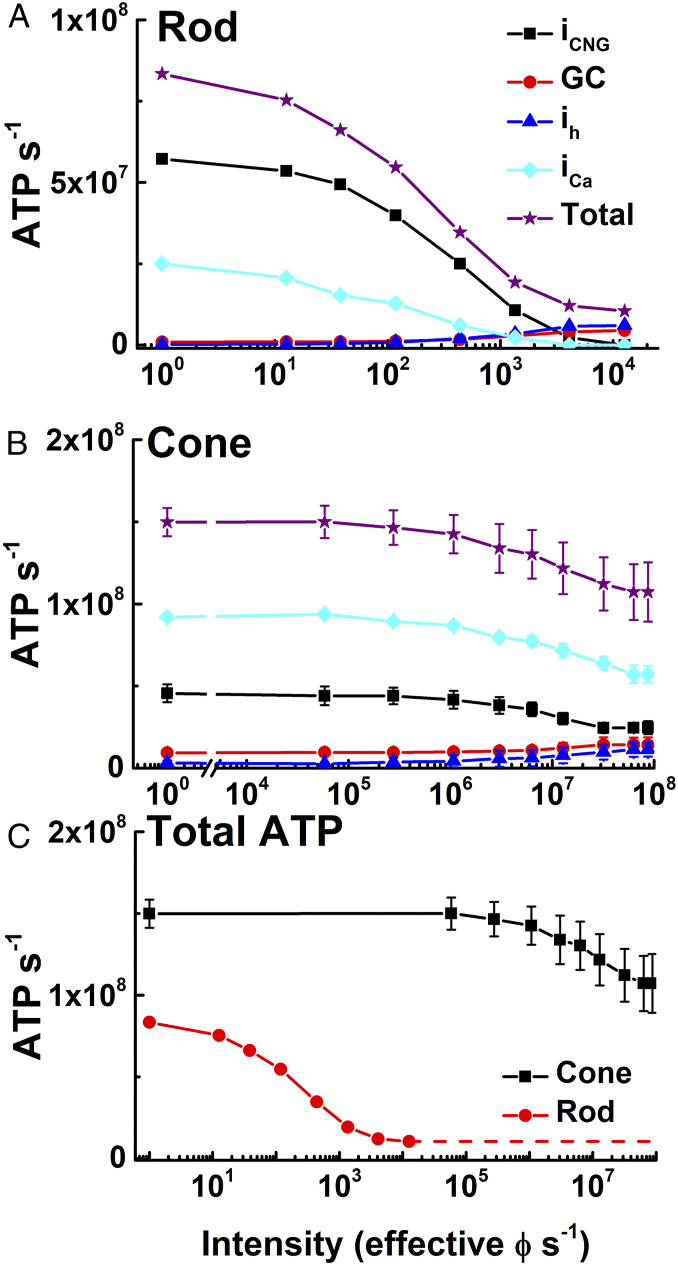
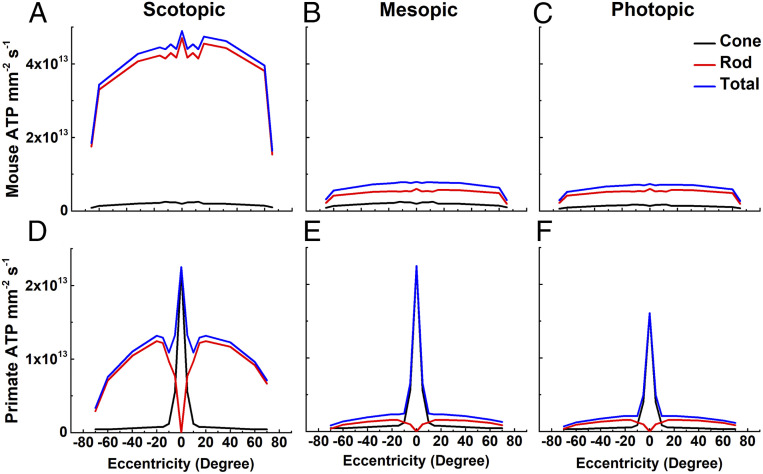
We have used recent measurements of mammalian cone light responses and voltage-gated currents to calculate cone ATP utilization and compare it to that of rods. The largest expenditure of ATP results from ion transport, particularly from removal of Na+ entering outer segment light-dependent channels and inner segment hyperpolarization-activated cyclic nucleotide-gated channels, and from ATP-dependent pumping of Ca2+ entering voltage-gated channels at the synaptic terminal. Single cones expend nearly twice as much energy as single rods in darkness, largely because they make more synapses with second-order retinal cells and thus must extrude more Ca2+ In daylight, cone ATP utilization per cell remains high because cones never remain saturated and must continue to export Na+ and synaptic Ca2+ even in bright illumination. In mouse and human retina, rods greatly outnumber cones and consume more energy overall even in background light. In primates, however, the high density of cones in the fovea produces a pronounced peak of ATP utilization, which becomes particularly prominent in daylight and may make this part of the retina especially sensitive to changes in energy availability.
Keywords: degeneration; fovea; metabolism; photoreceptor; retina.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Attwell D., Laughlin S. B., An energy budget for signaling in the grey matter of the brain. J. Cereb. Blood Flow Metab. 21, 1133–1145 (2001). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
