

Structures of capsid and capsid-associated tegument complex inside the Epstein-Barr virus
- PMID: 32719506
- PMCID: PMC7546529
- DOI: 10.1038/s41564-020-0758-1
Structures of capsid and capsid-associated tegument complex inside the Epstein-Barr virus
Abstract
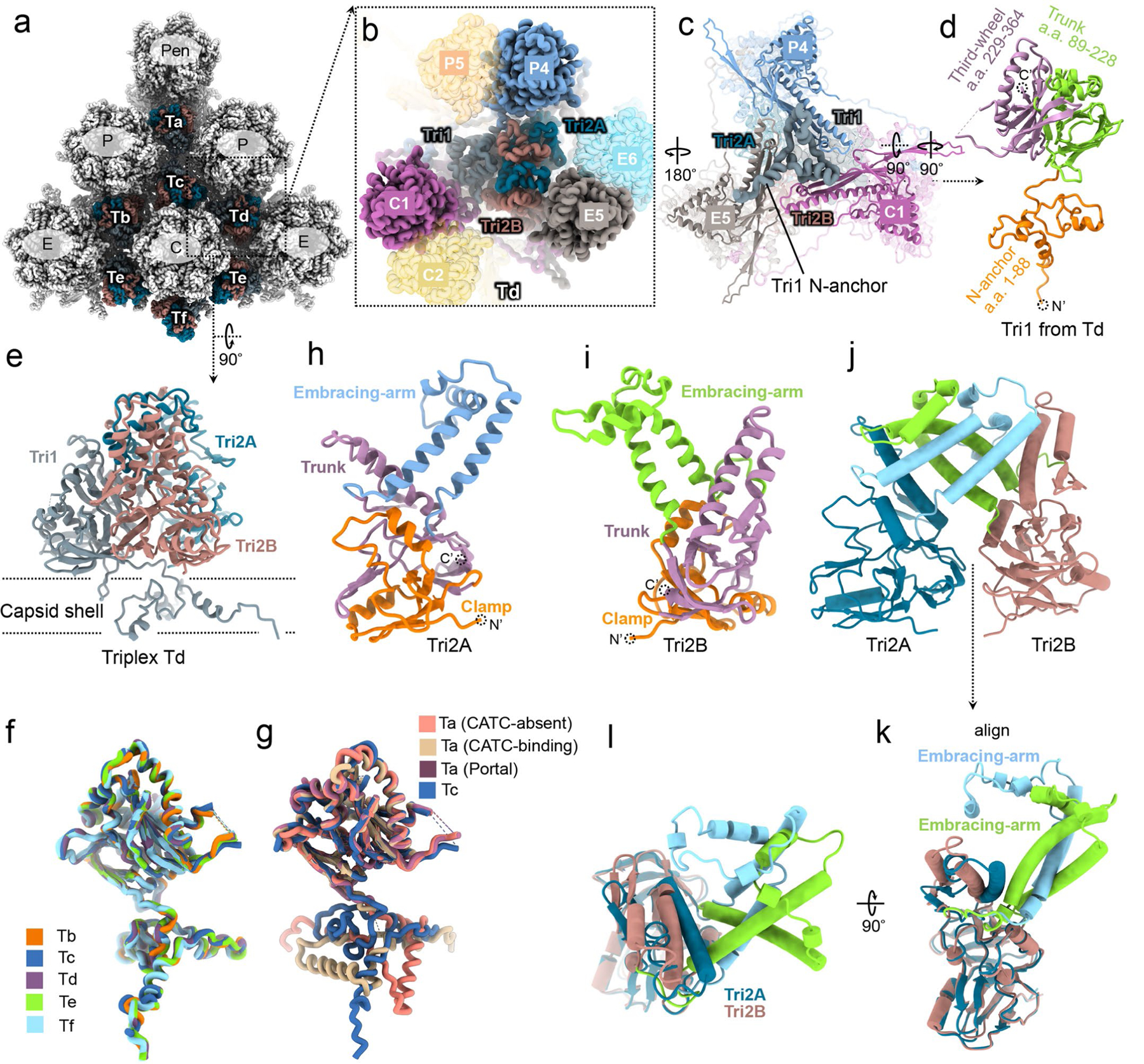
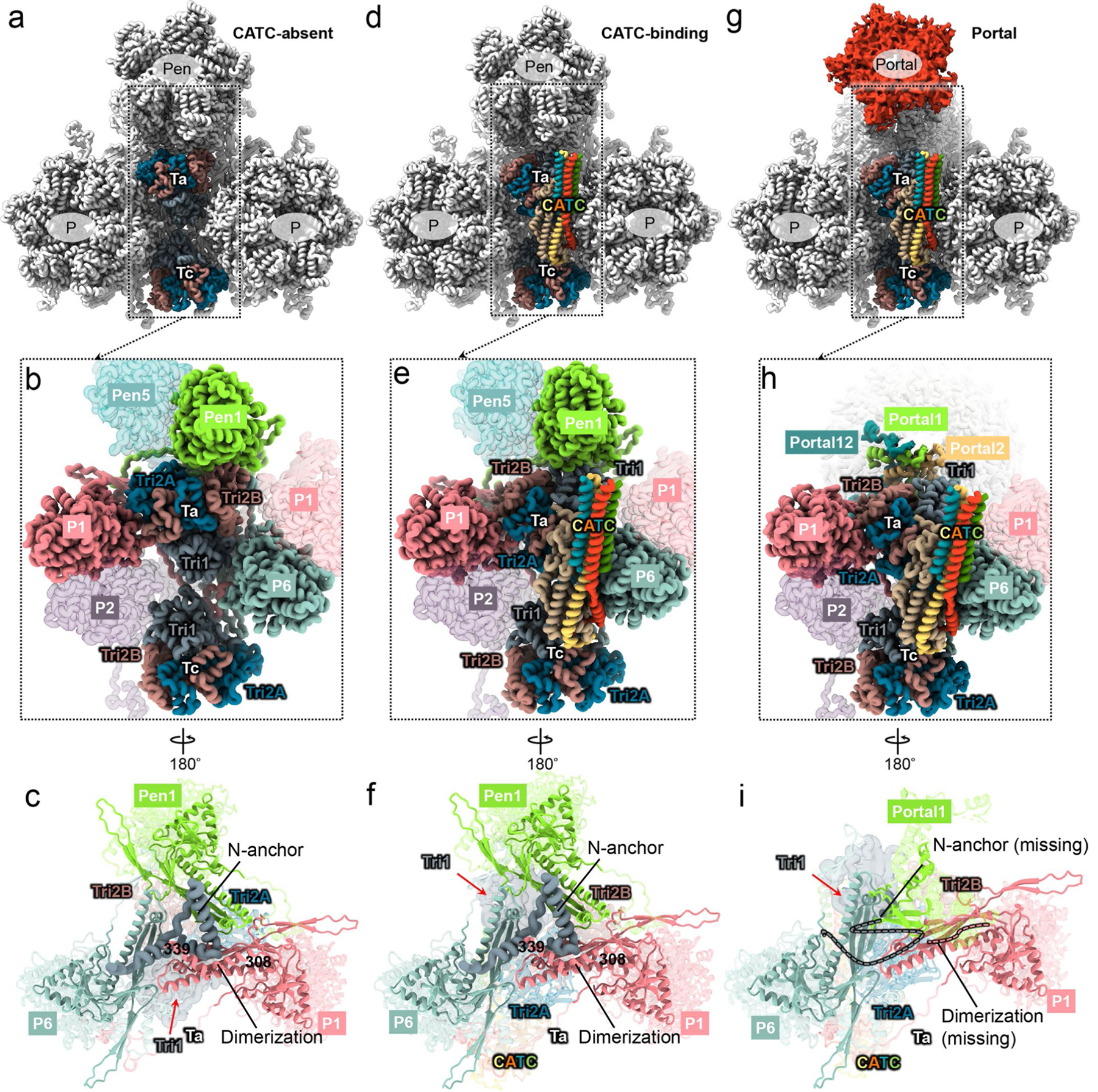
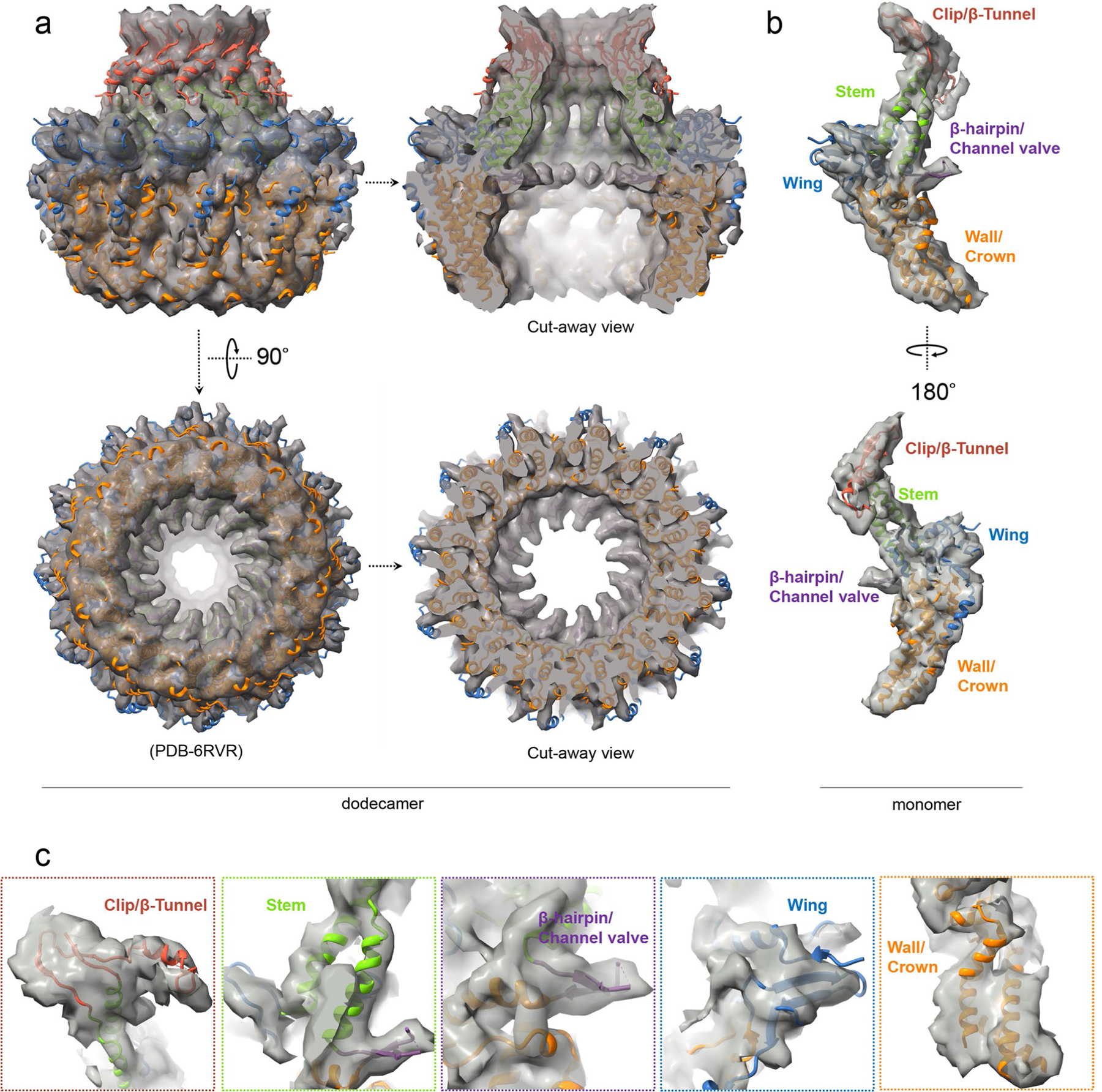
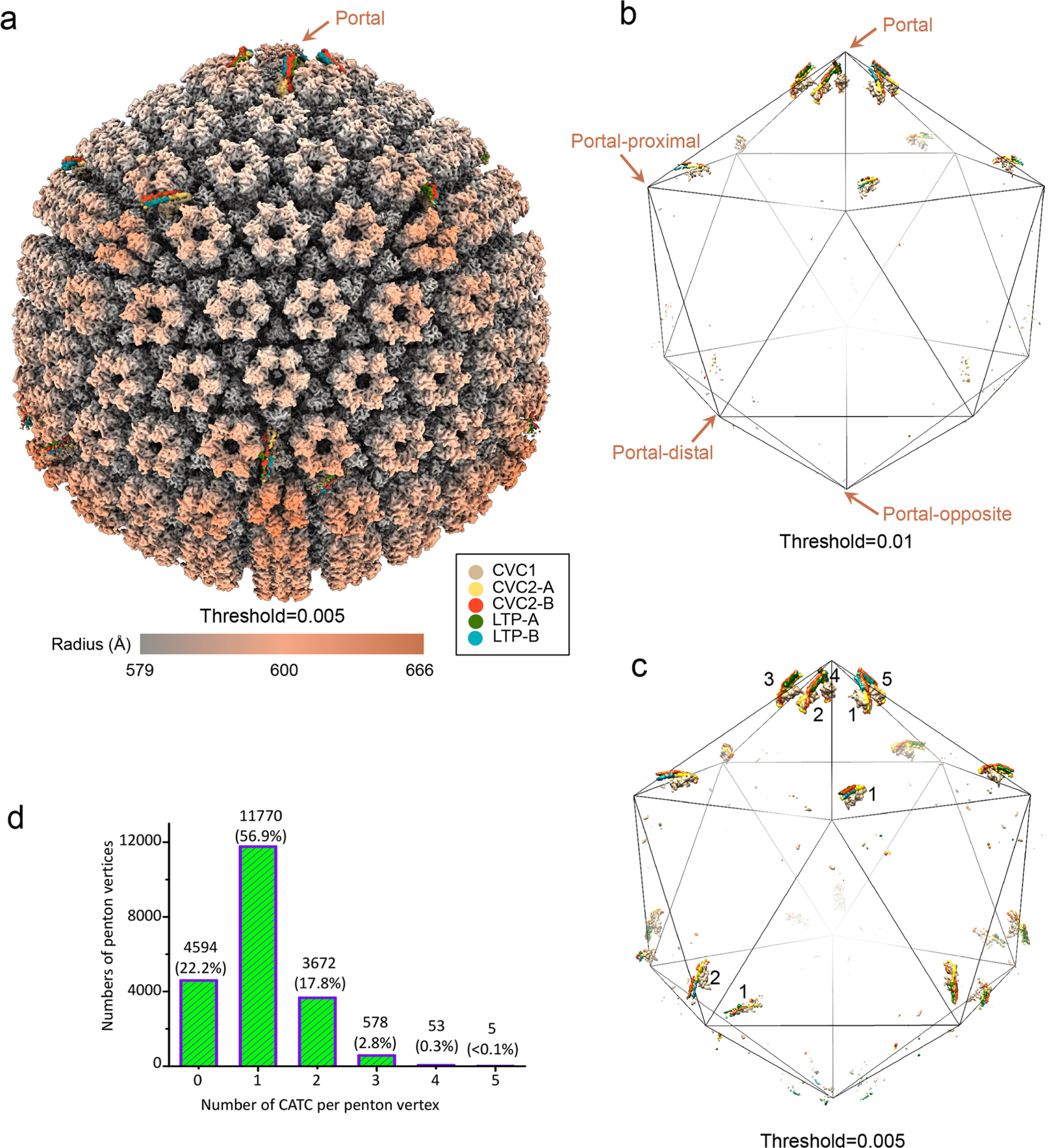
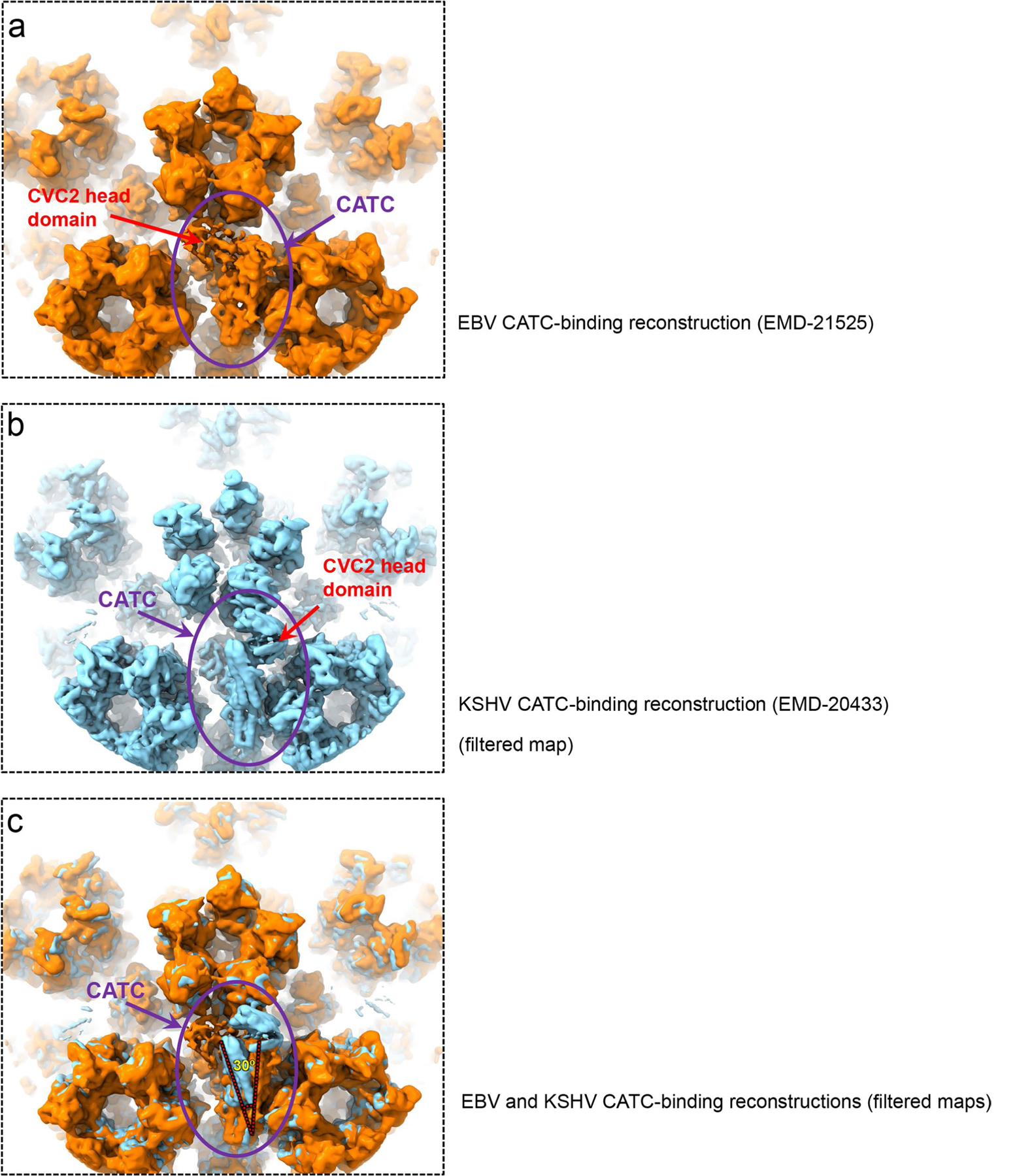
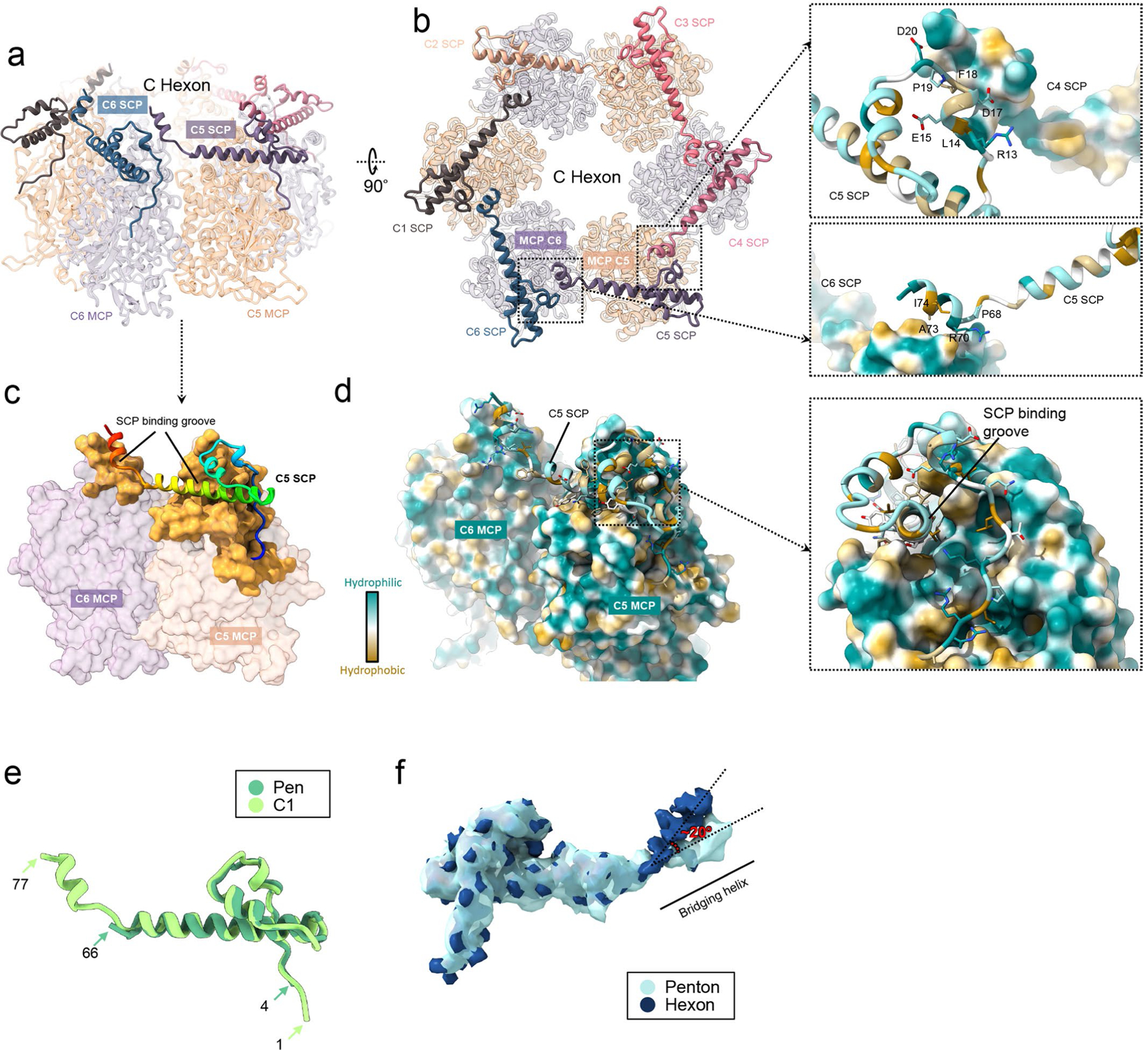
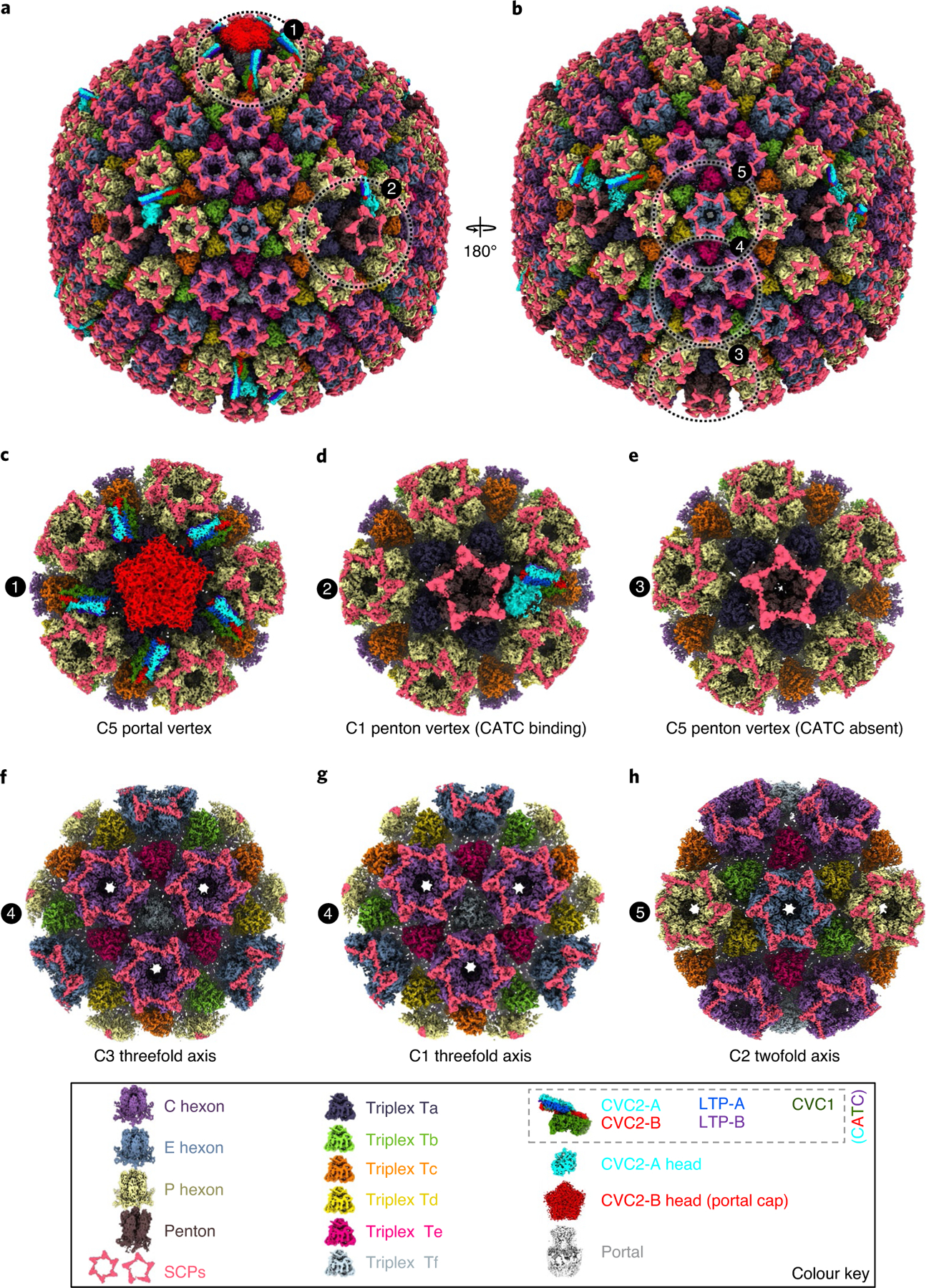
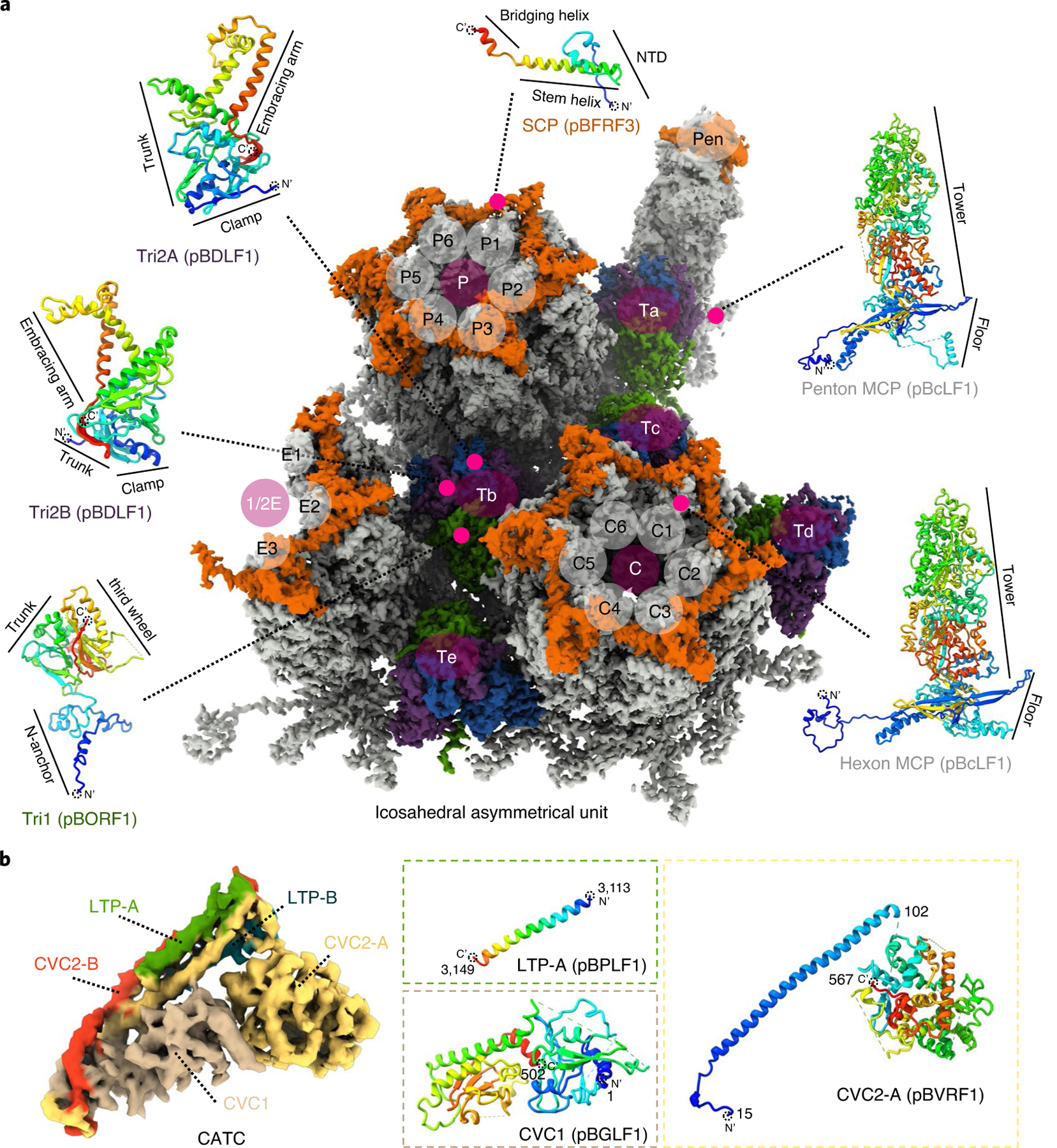
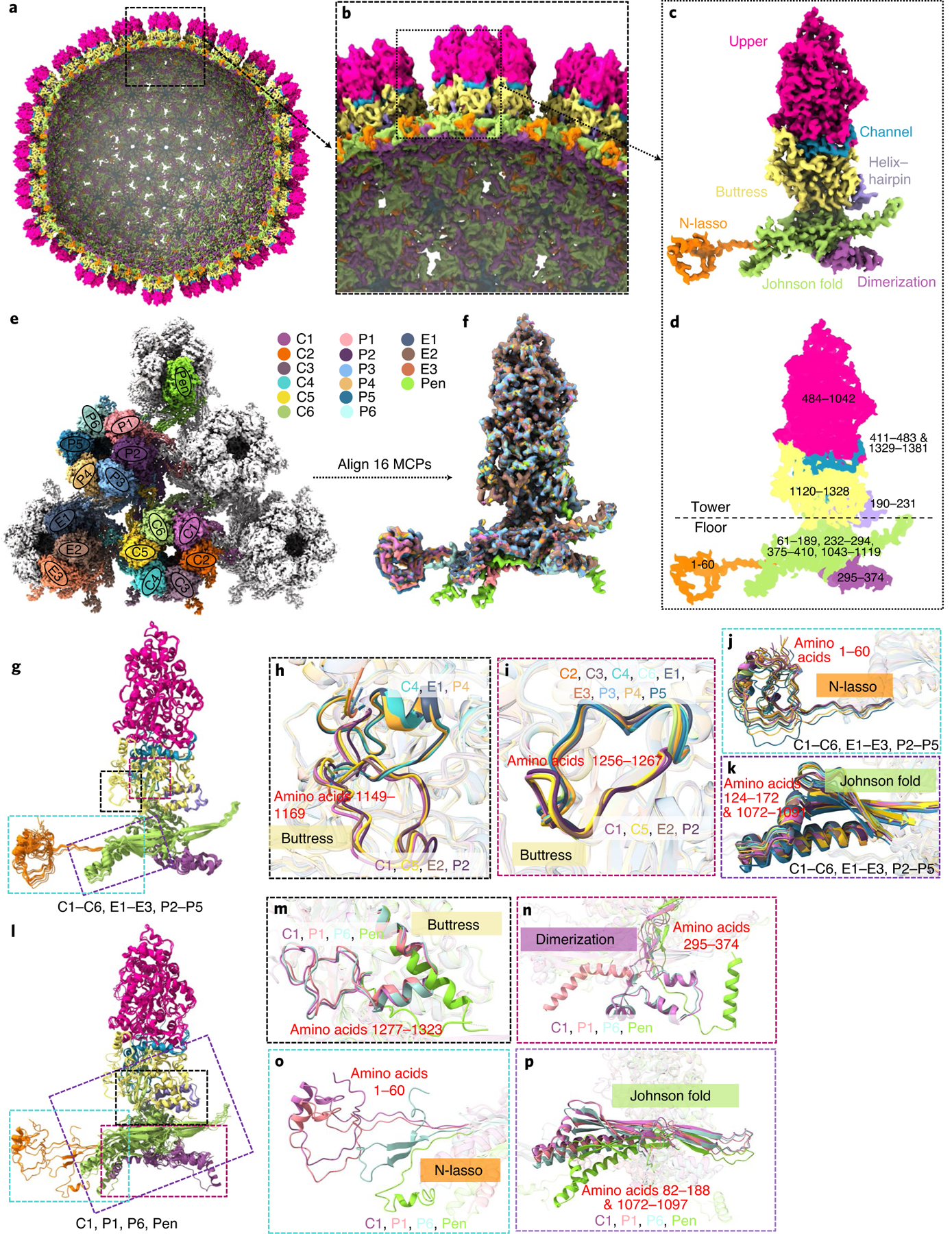
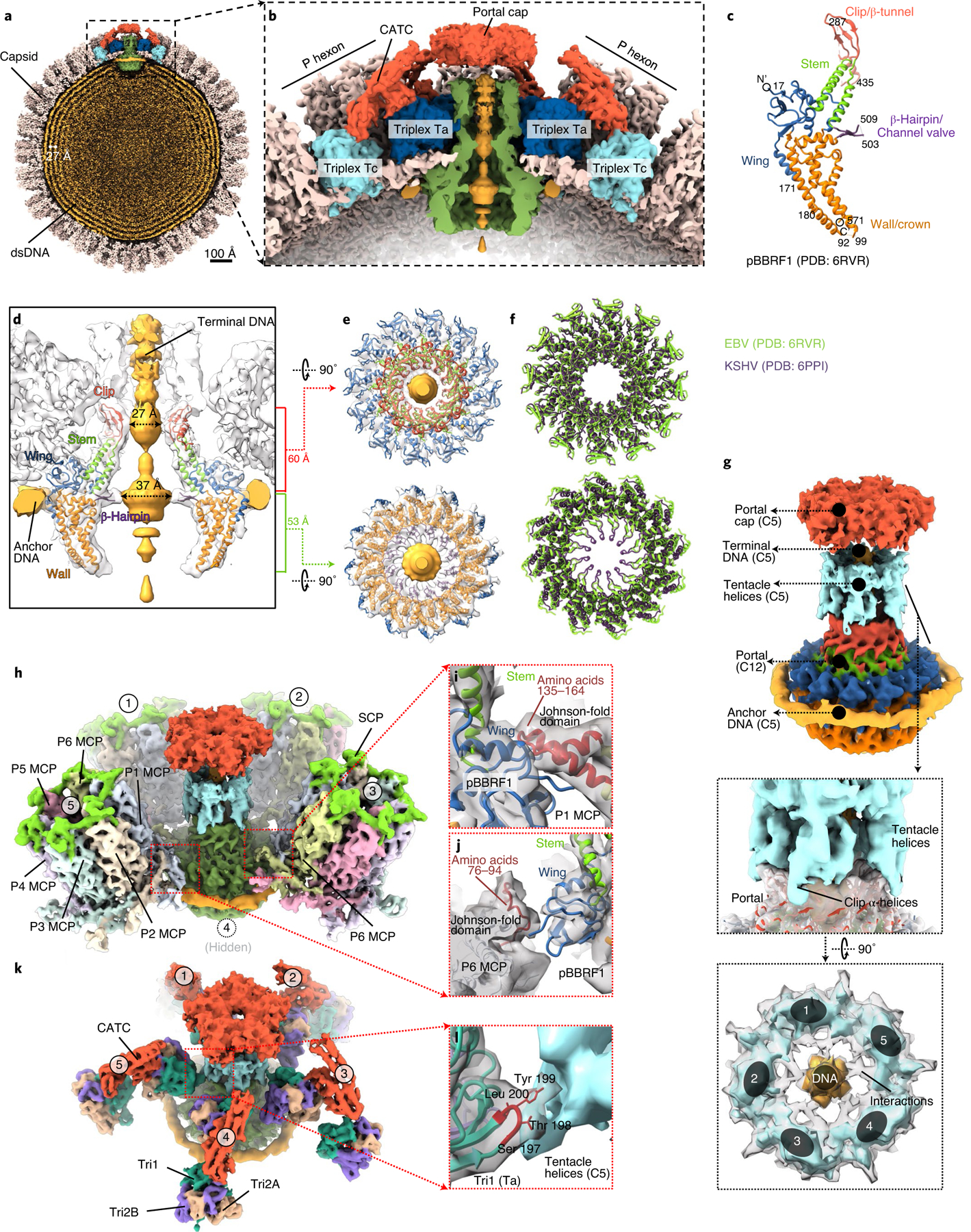
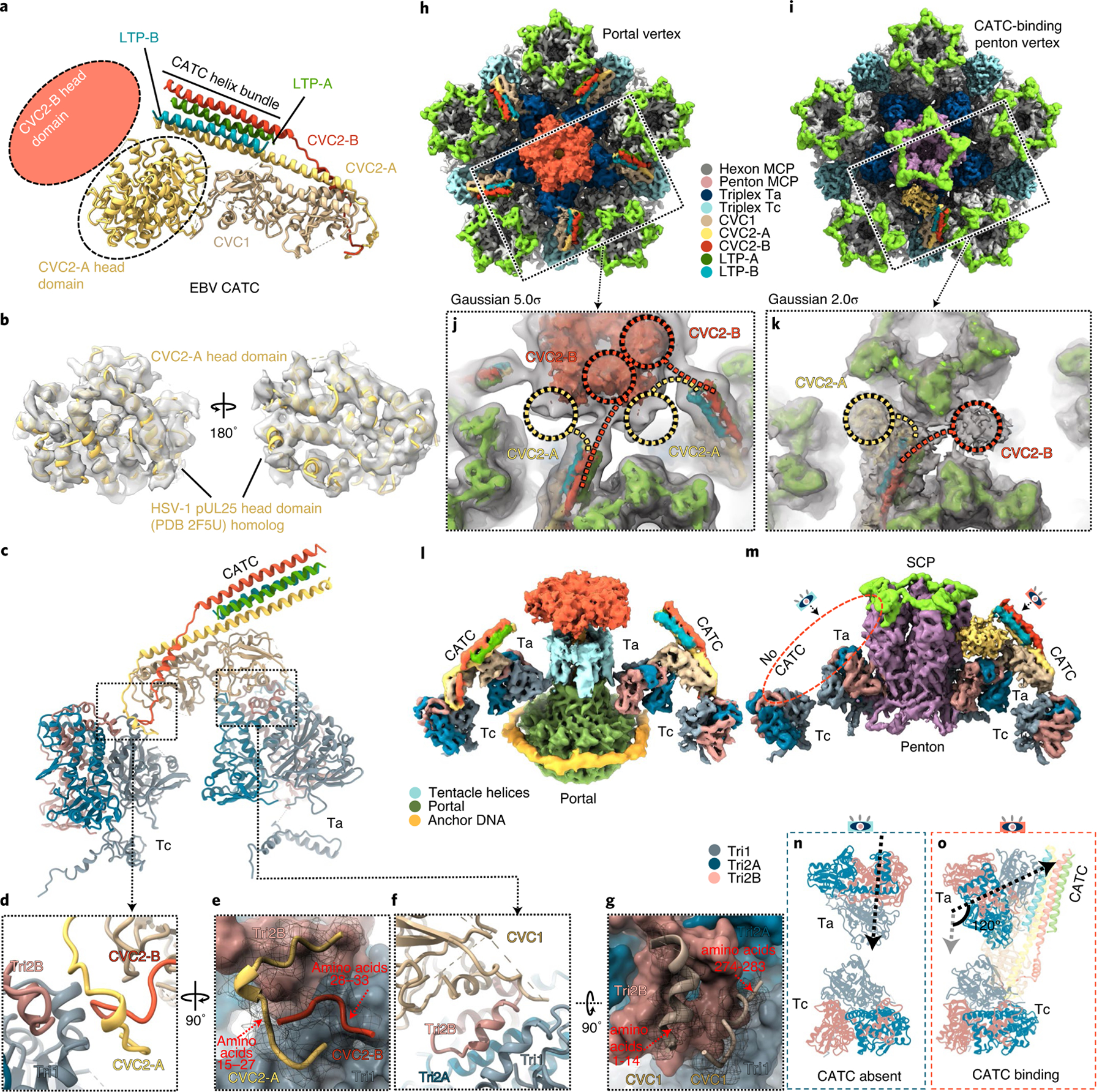
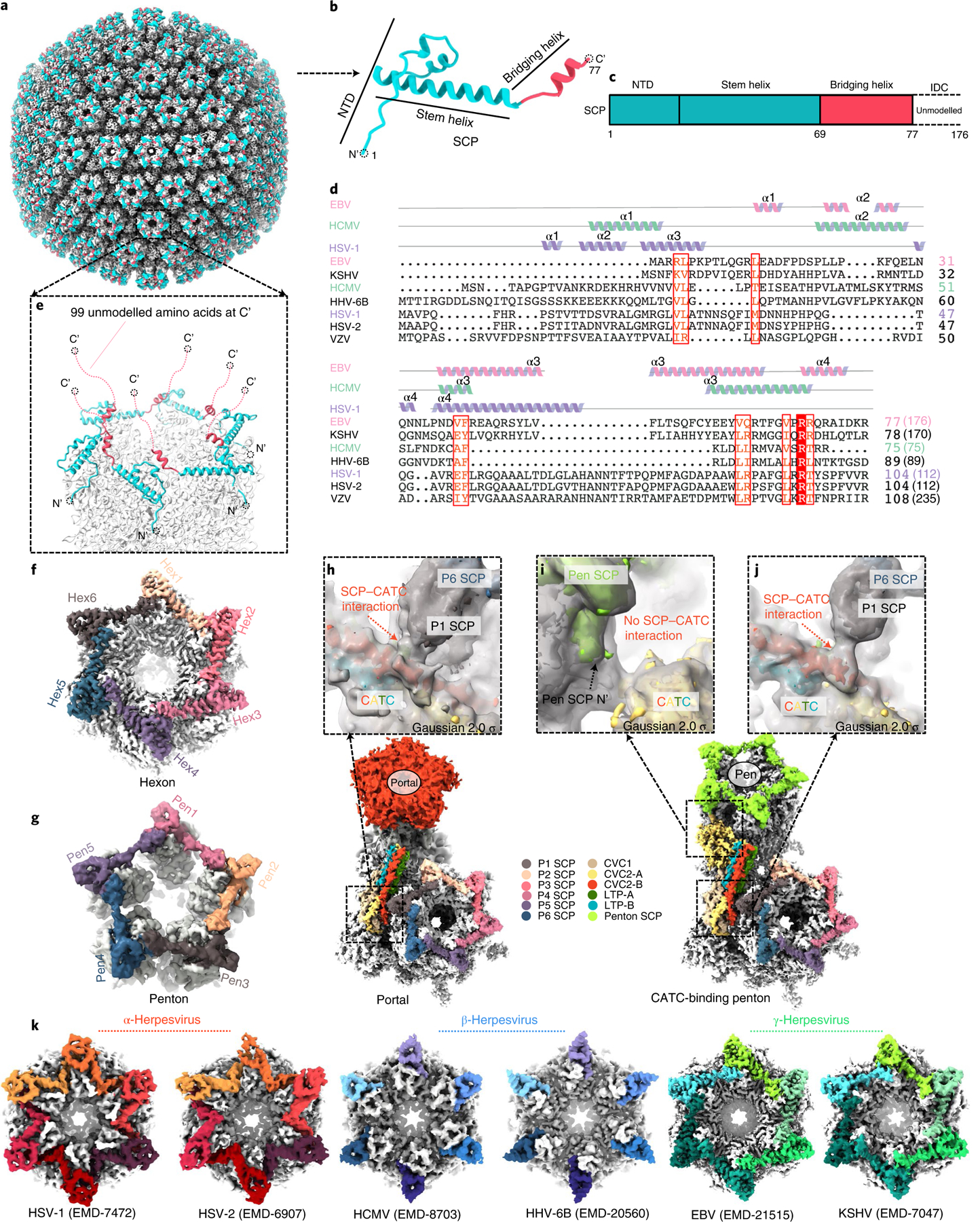
As the first discovered human cancer virus, Epstein-Barr virus (EBV) causes Burkitt's lymphoma and nasopharyngeal carcinoma. Isolating virions for determining high-resolution structures has been hindered by latency-a hallmark of EBV infection-and atomic structures are thus available only for recombinantly expressed EBV proteins. In the present study, by symmetry relaxation and subparticle reconstruction, we have determined near-atomic-resolution structures of the EBV capsid with an asymmetrically attached DNA-translocating portal and capsid-associated tegument complexes from cryogenic electron microscopy images of just 2,048 EBV virions obtained by chemical induction. The resulting atomic models reveal structural plasticity among the 20 conformers of the major capsid protein, 2 conformers of the small capsid protein (SCP), 4 conformers of the triplex monomer proteins and 2 conformers of the triplex dimer proteins. Plasticity reaches the greatest level at the capsid-tegument interfaces involving SCP and capsid-associated tegument complexes (CATC): SCPs crown pentons/hexons and mediate tegument protein binding, and CATCs bind and rotate all five periportal triplexes, but notably only about one peri-penton triplex. These results offer insights into the EBV capsid assembly and a mechanism for recruiting cell-regulating factors into the tegument compartment as 'cargoes', and should inform future anti-EBV strategies.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
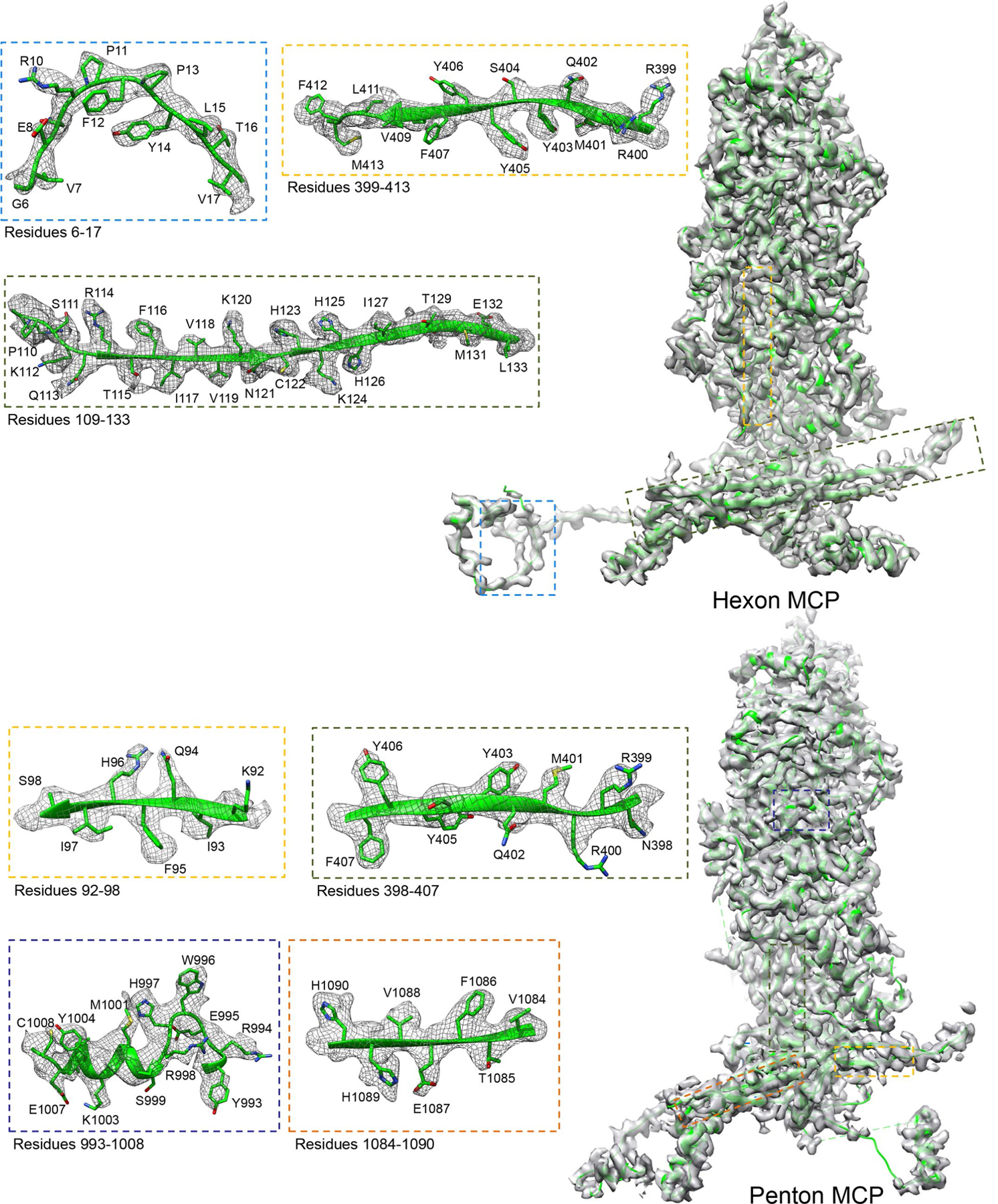
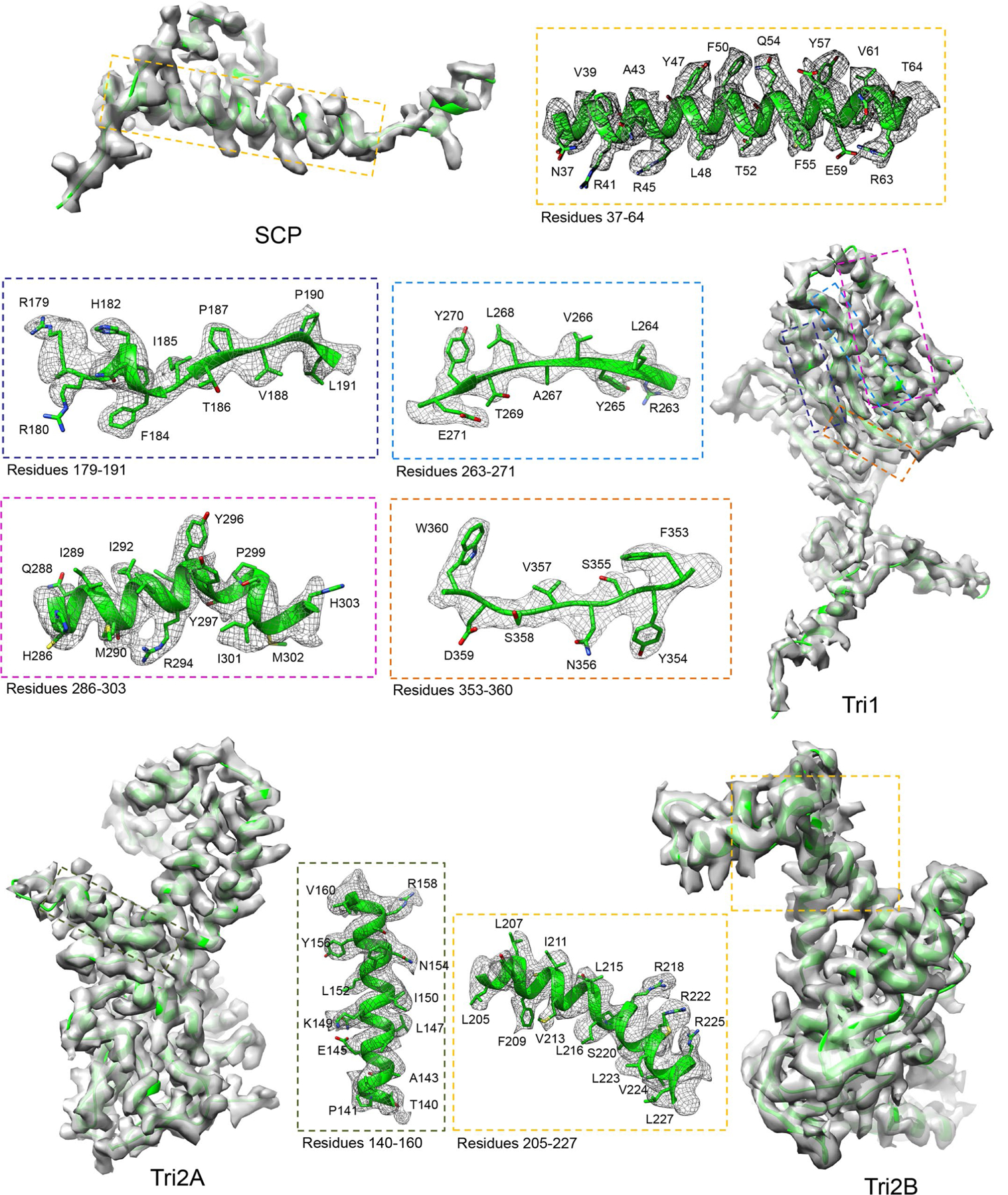
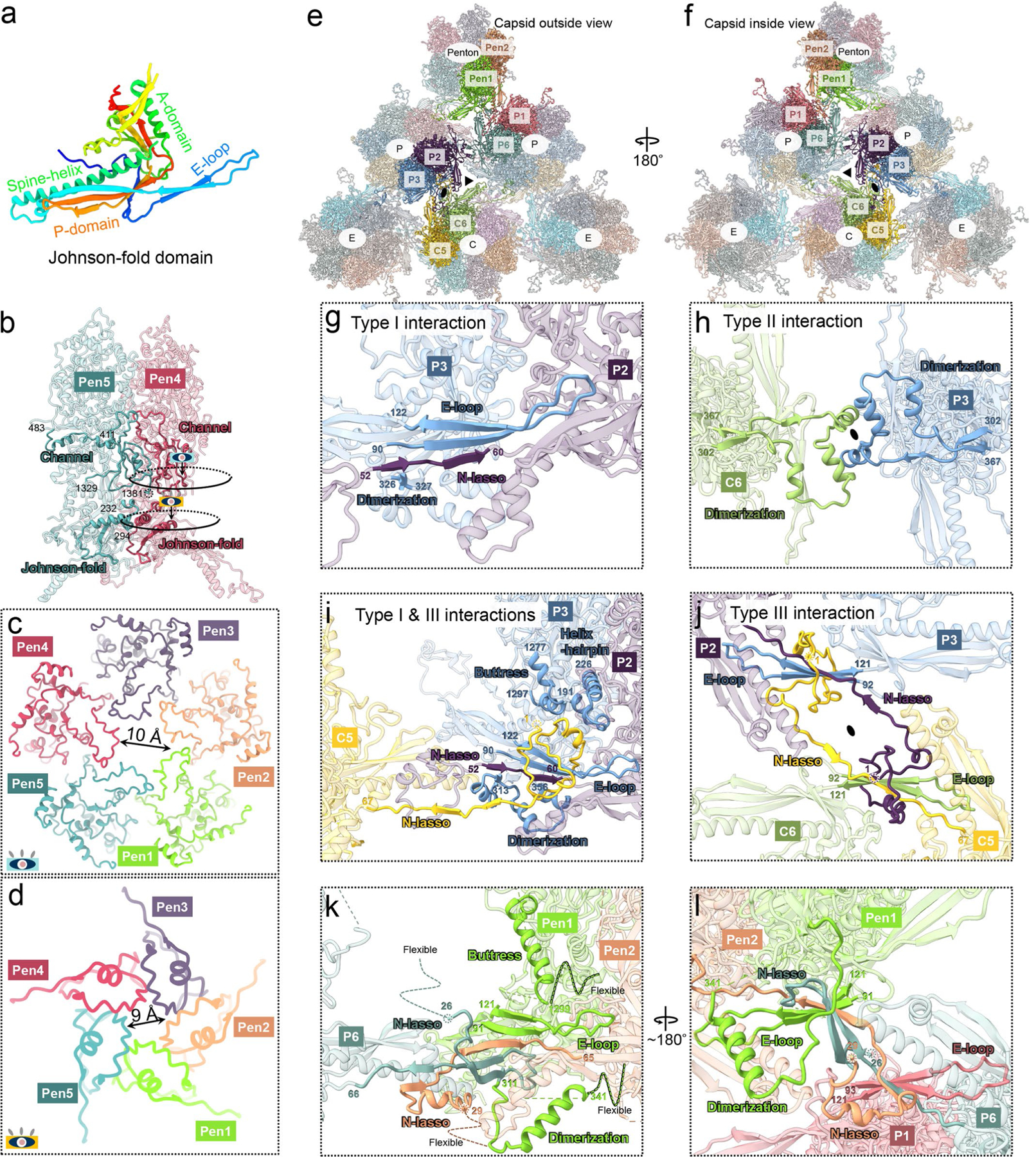
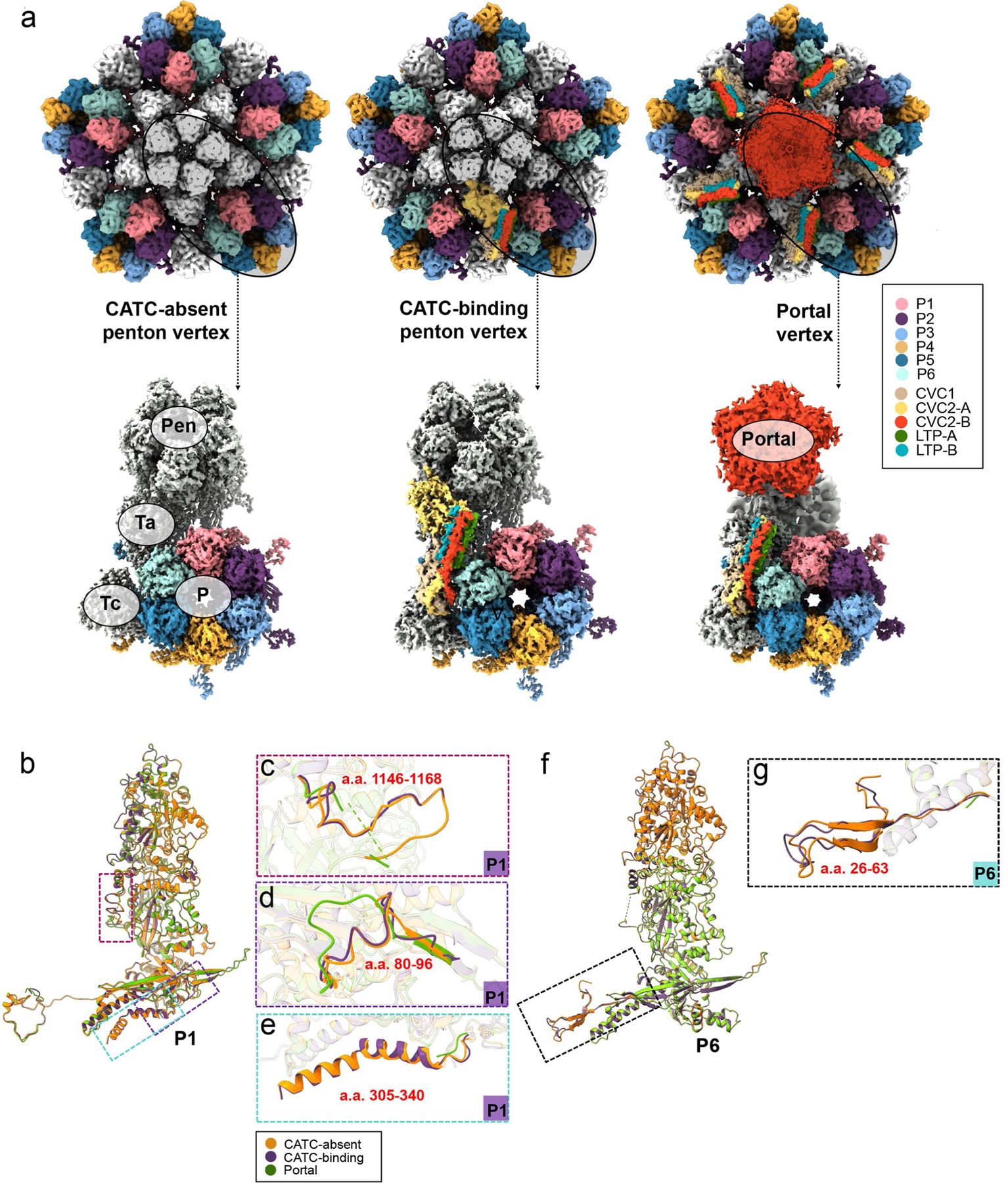
References
-
- Epstein MA, Achong BG & Barr YM Virus particles in cultured lymphoblasts from Burkitt’s lymphoma. Lancet 283, 702–703 (1964). - PubMed
-
- Salahuddin SZ et al. Isolation of a new virus, HBLV, in patients with lymphoproliferative disorders. Science 234, 596–601 (1986). - PubMed
-
- Andersson JP Clinical aspects on Epstein–Barr virus infection. Scand. J. Infect. Dis. Suppl 80, 94–104 (1991). - PubMed
-
- Bochkarev A et al. Crystal structure of the DNA-binding domain of the Epstein–Barr virus origin-binding protein EBNA1. Cell 83, 39–46 (1995). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
