

Dihydroxyacetone phosphate signals glucose availability to mTORC1
- PMID: 32719541
- PMCID: PMC7995735
- DOI: 10.1038/s42255-020-0250-5
Dihydroxyacetone phosphate signals glucose availability to mTORC1
Abstract
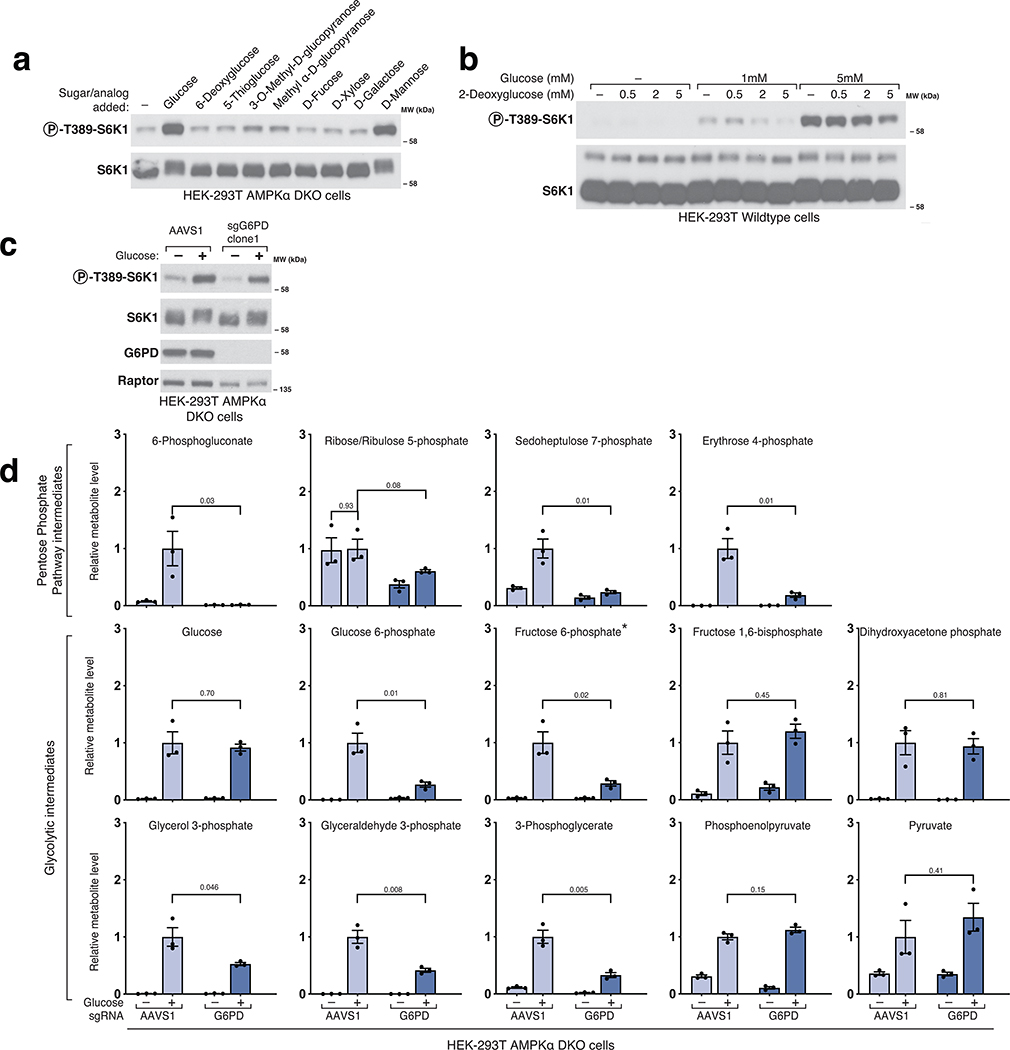
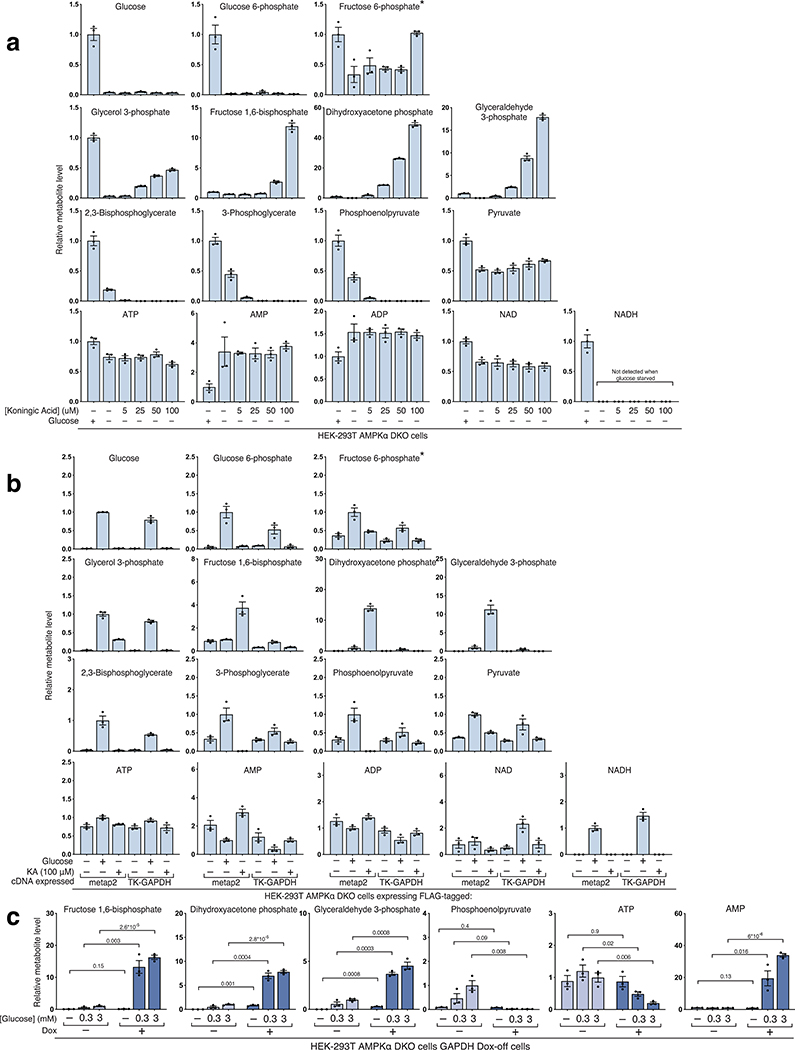
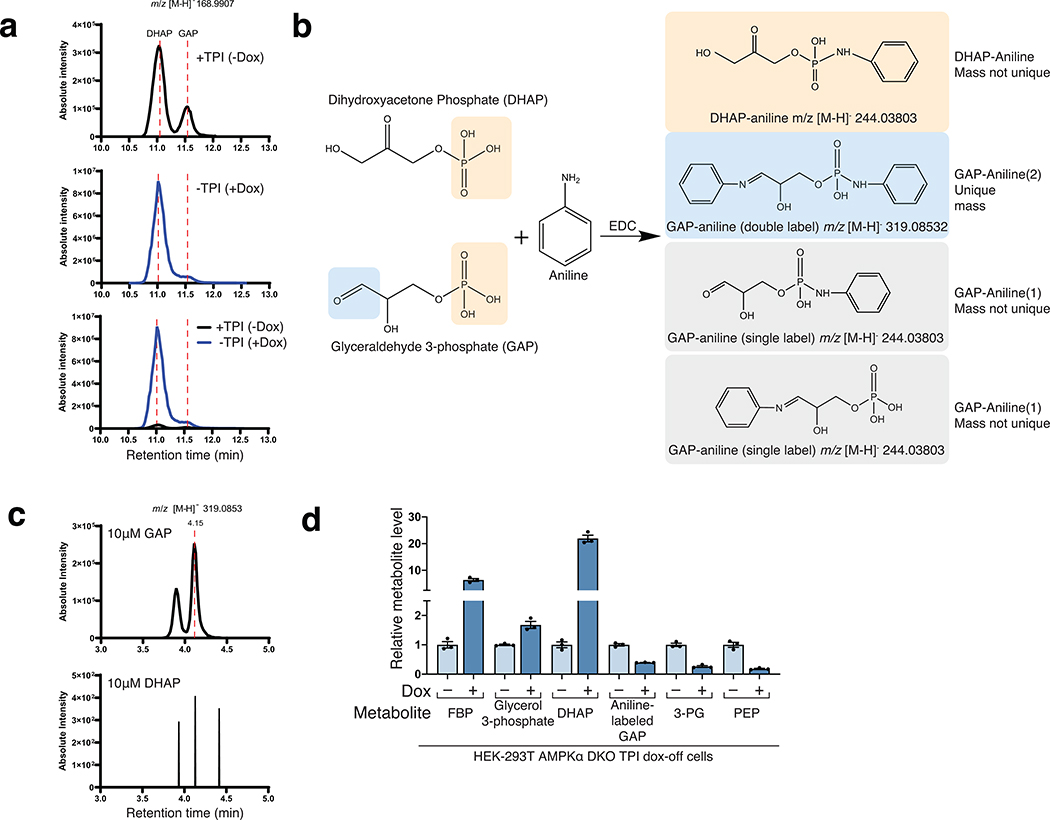
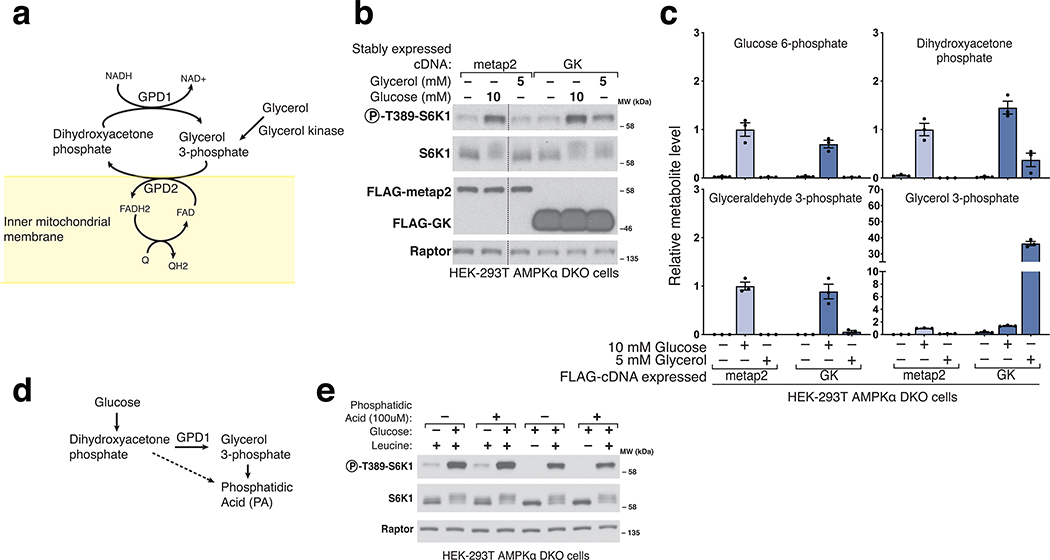
The mechanistic target of rapamycin complex 1 (mTORC1) kinase regulates cell growth by setting the balance between anabolic and catabolic processes. To be active, mTORC1 requires the environmental presence of amino acids and glucose. While a mechanistic understanding of amino acid sensing by mTORC1 is emerging, how glucose activates mTORC1 remains mysterious. Here, we used metabolically engineered human cells lacking the canonical energy sensor AMP-activated protein kinase to identify glucose-derived metabolites required to activate mTORC1 independent of energetic stress. We show that mTORC1 senses a metabolite downstream of the aldolase and upstream of the GAPDH-catalysed steps of glycolysis and pinpoint dihydroxyacetone phosphate (DHAP) as the key molecule. In cells expressing a triose kinase, the synthesis of DHAP from DHA is sufficient to activate mTORC1 even in the absence of glucose. DHAP is a precursor for lipid synthesis, a process under the control of mTORC1, which provides a potential rationale for the sensing of DHAP by mTORC1.
Conflict of interest statement
Competing Interests Statement
DMS is a founder, shareholder, and a member of the scientific advisory board for Navitor Pharmaceuticals, which is targeting the mTORC1 pathway for therapeutic benefit. J.M.O. is a shareholder of Navitor Pharmaceuticals. The other authors declare no competing intersts.
Figures
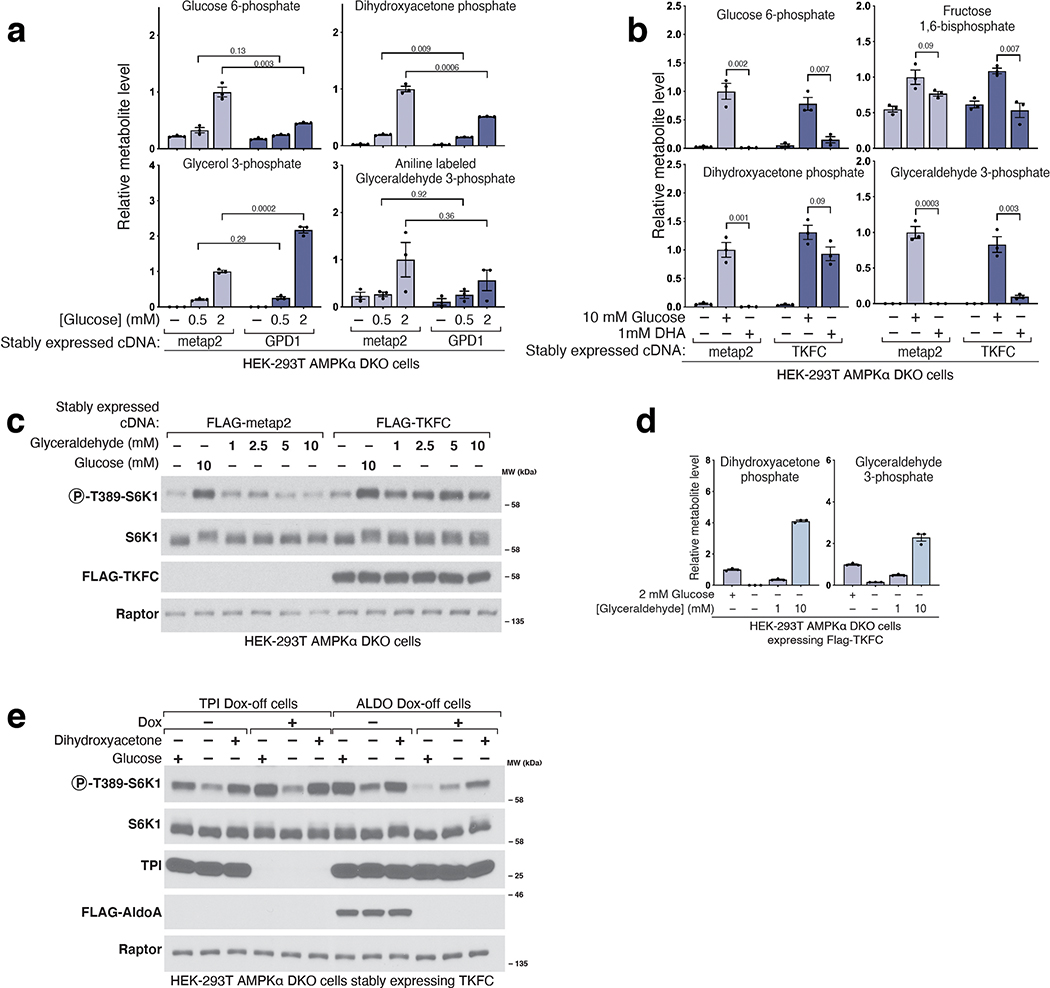
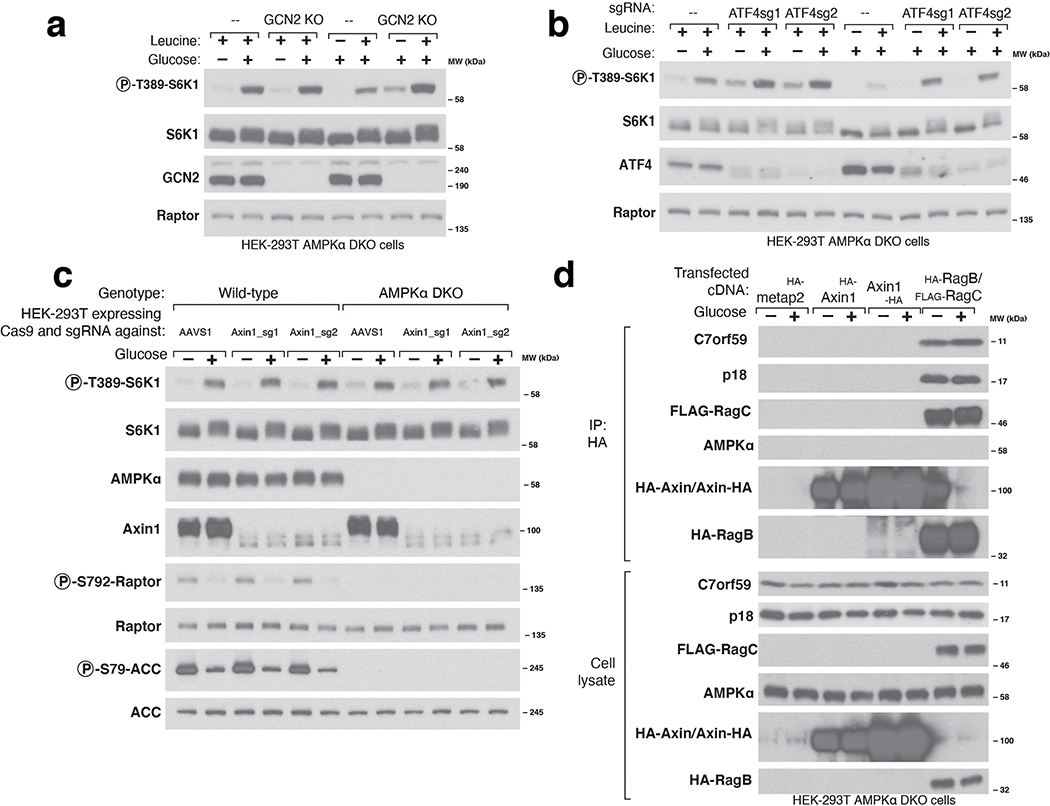
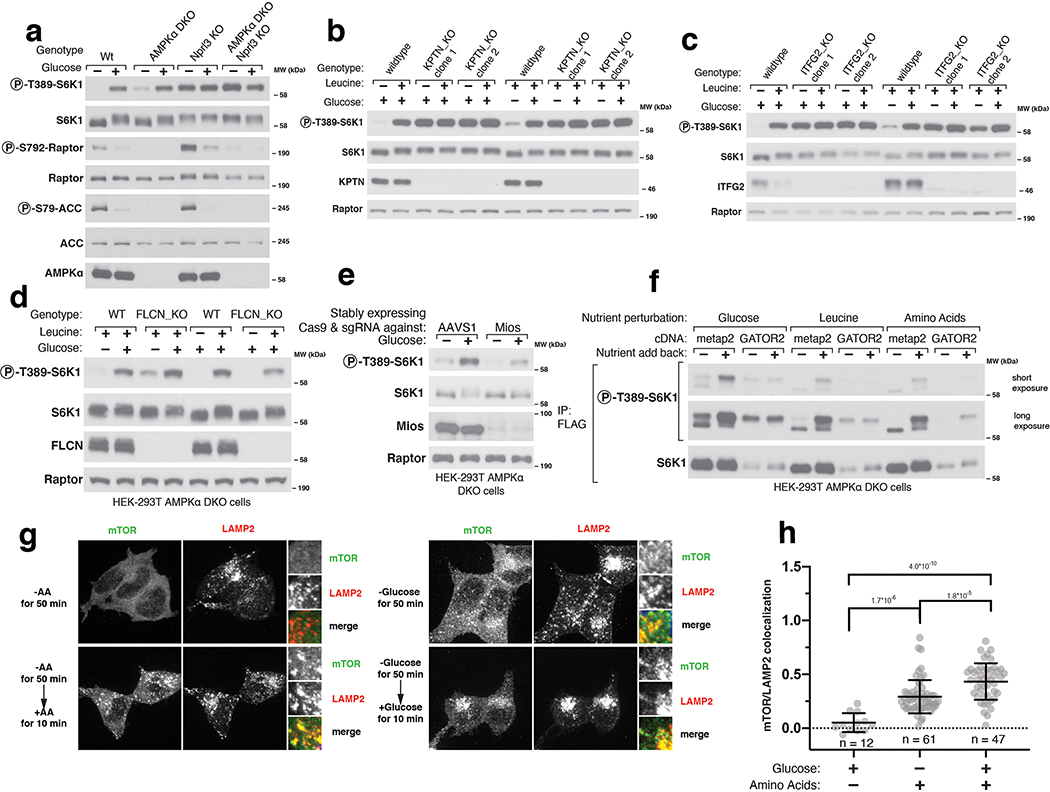
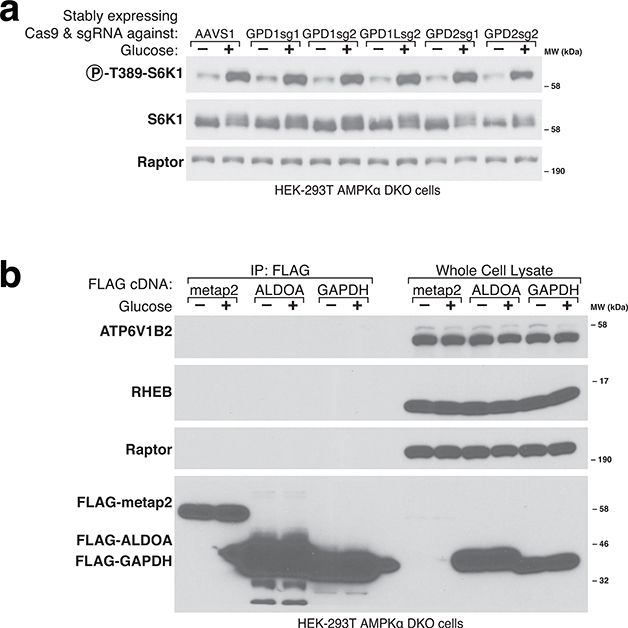
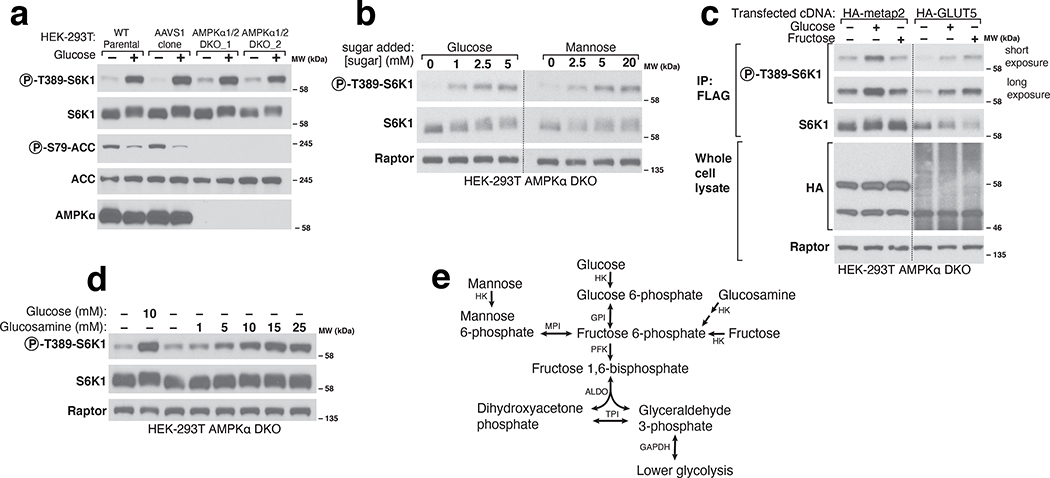
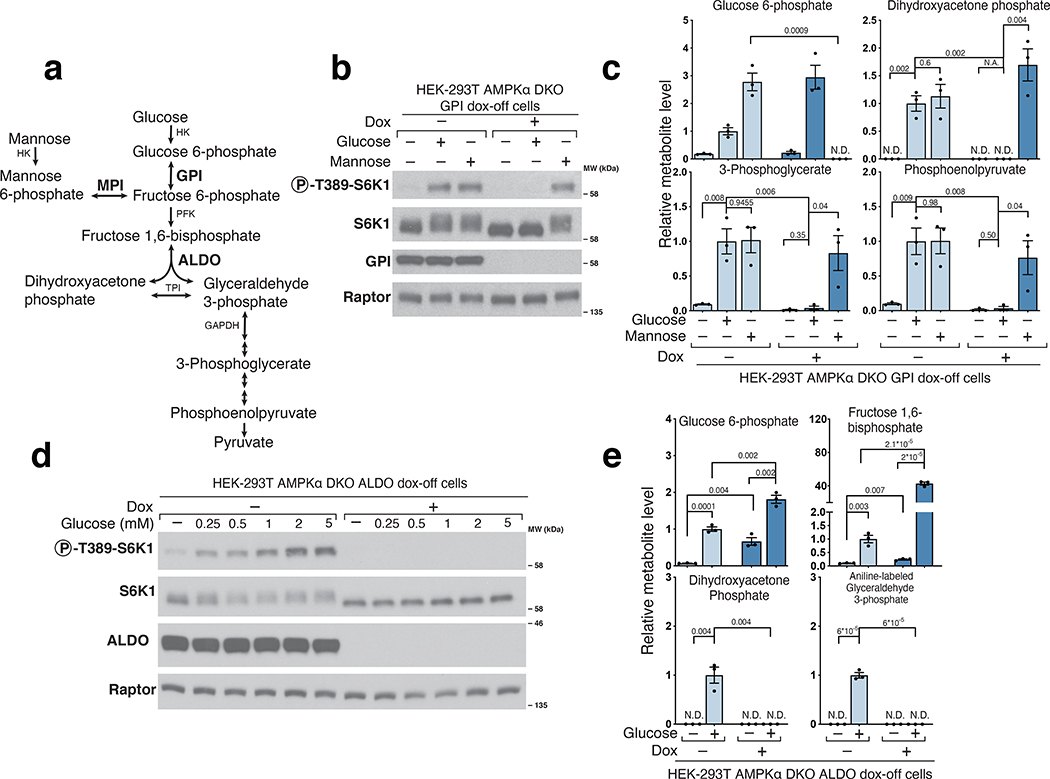
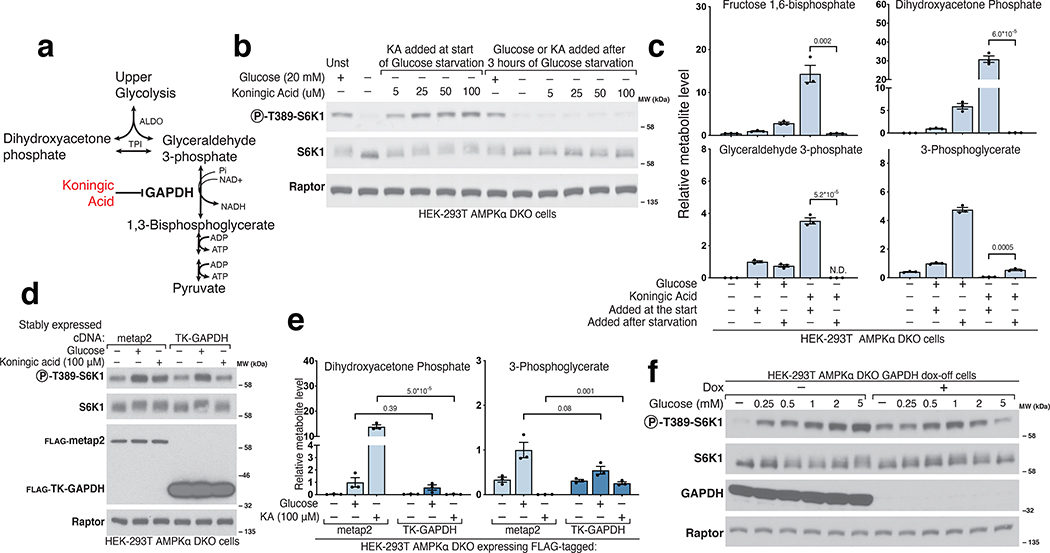
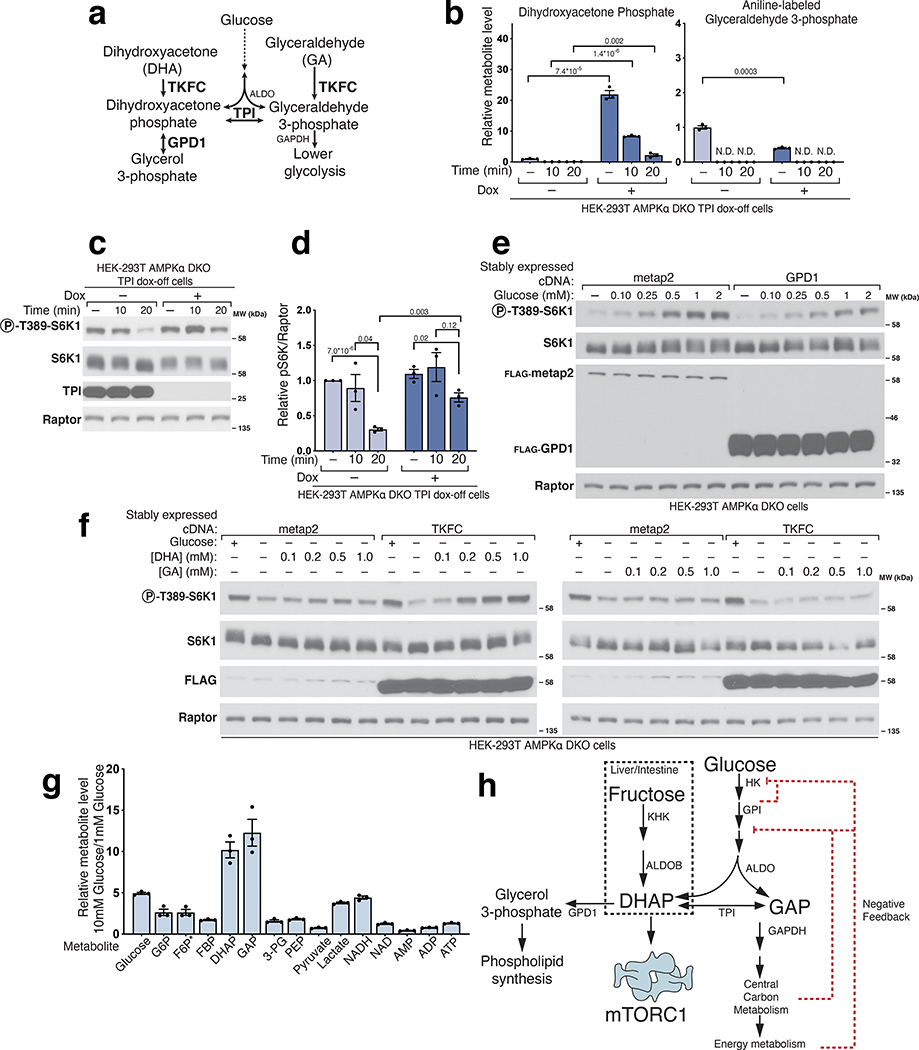
References
-
- Kubrusly MS et al. A role for mammalian target of rapamycin (mTOR) pathway in non alcoholic steatohepatitis related-cirrhosis. Histol. Histopathol 25, 1123–1131 (2010). - PubMed
-
- Ardestani A, Lupse B, Kido Y, Leibowitz G & Maedler K mTORC1 Signaling: A Double-Edged Sword in Diabetic β Cells. Cell Metab 27, 314–331 (2018). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
