

Transcriptional interference at tandem lncRNA and protein-coding genes: an emerging theme in regulation of cellular nutrient homeostasis
- PMID: 32720681
- PMCID: PMC7470944
- DOI: 10.1093/nar/gkaa630
Transcriptional interference at tandem lncRNA and protein-coding genes: an emerging theme in regulation of cellular nutrient homeostasis
Abstract
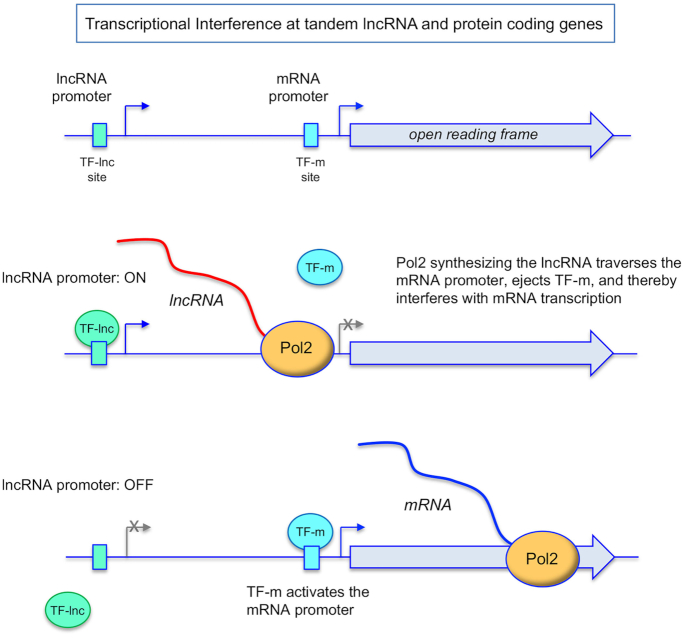
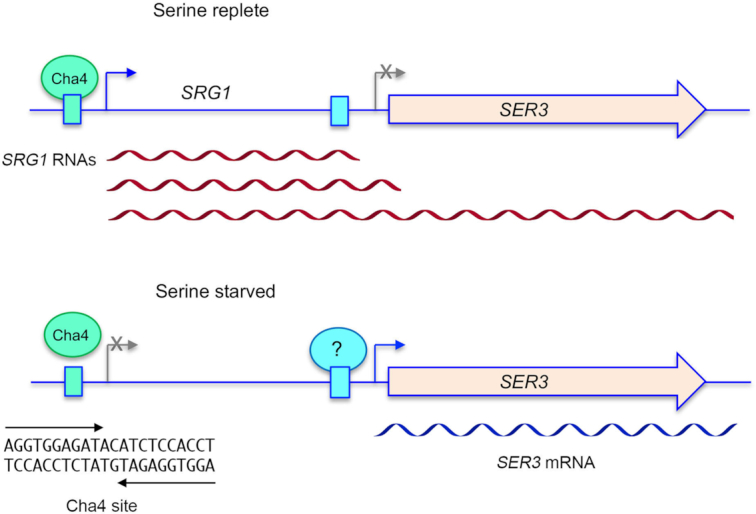
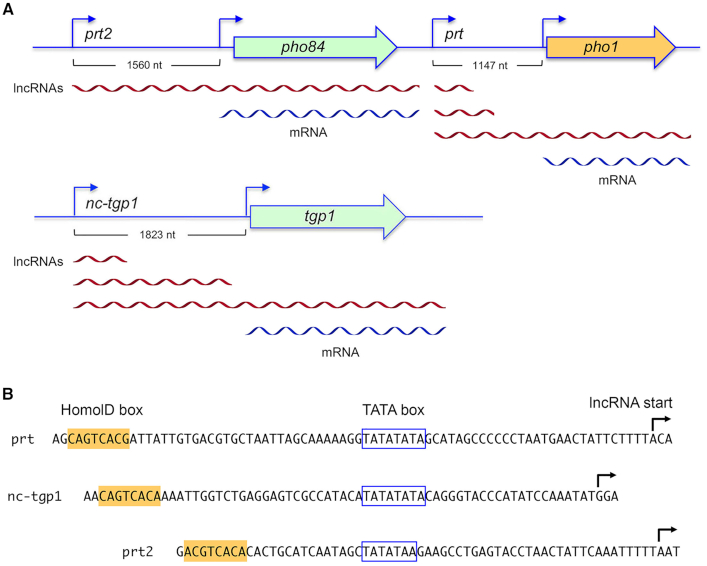
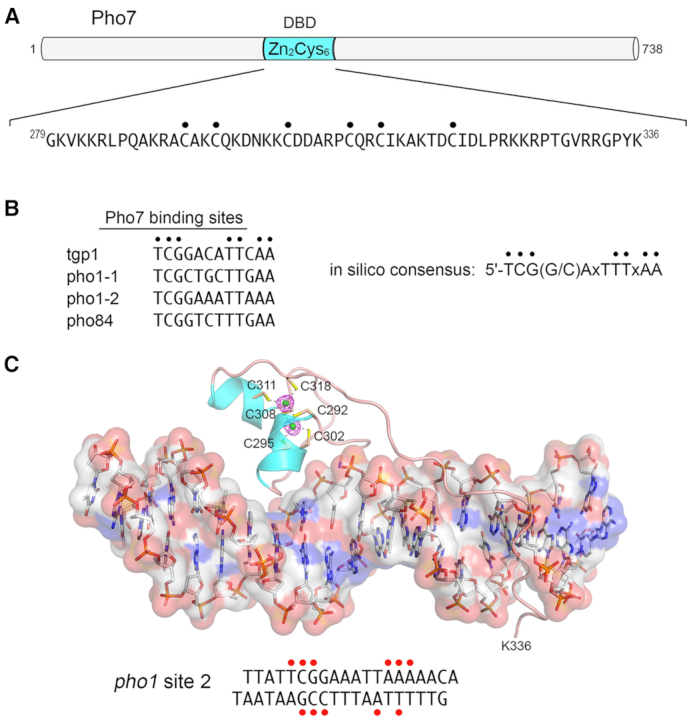
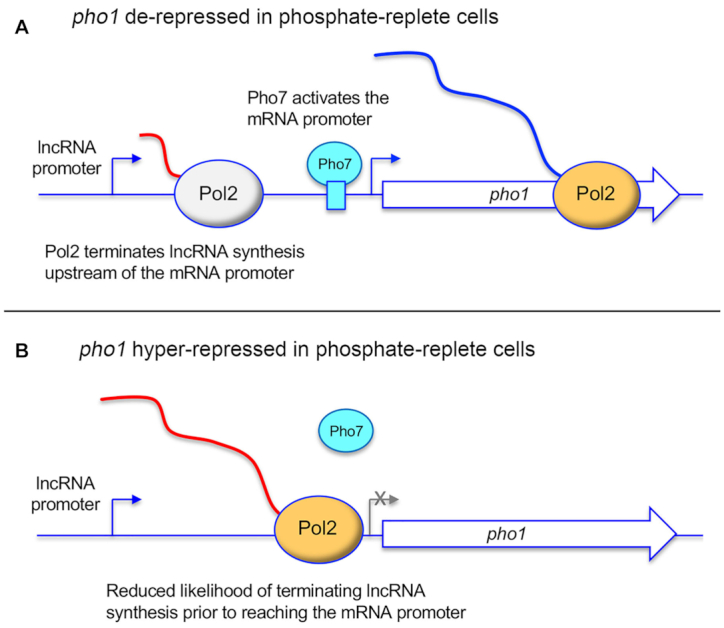
Tandem transcription interference occurs when the act of transcription from an upstream promoter suppresses utilization of a co-oriented downstream promoter. Because eukaryal genomes are liberally interspersed with transcription units specifying long non-coding (lnc) RNAs, there are many opportunities for lncRNA synthesis to negatively affect a neighboring protein-coding gene. Here, I review two eukaryal systems in which lncRNA interference with mRNA expression underlies a regulated biological response to nutrient availability. Budding yeast SER3 is repressed under serine-replete conditions by transcription of an upstream SRG1 lncRNA that traverses the SER3 promoter and elicits occlusive nucleosome rearrangements. SER3 is de-repressed by serine withdrawal, which leads to shut-off of SRG1 synthesis. The fission yeast phosphate homeostasis (PHO) regulon comprises three phosphate acquisition genes - pho1, pho84, and tgp1 - that are repressed under phosphate-replete conditions by 5' flanking lncRNAs prt, prt2, and nc-tgp1, respectively. lncRNA transcription across the PHO mRNA promoters displaces activating transcription factor Pho7. PHO mRNAs are transcribed during phosphate starvation when lncRNA synthesis abates. The PHO regulon is de-repressed in phosphate-replete cells by genetic manipulations that favor 'precocious' lncRNA 3'-processing/termination upstream of the mRNA promoters. PHO lncRNA termination is governed by the Pol2 CTD code and is subject to metabolite control by inositol pyrophosphates.
© The Author(s) 2020. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Proudfoot N.J. Transcriptional interference and termination between duplicated α-globin gene constructs suggests a novel mechanism for gene regulation. Nature. 1986; 322:562–565. - PubMed
-
- Colin J., Candelli T., Porrua O., Boulay J., Zhu C., Lacroute F., Steinmetz L.M., Libri D.. Roadblock termination by Reb1p restricts cryptic and readthrough transcription. Mol. Cell. 2014; 56:667–680. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
