

CENP-A nucleosome-a chromatin-embedded pedestal for the centromere: lessons learned from structural biology
- PMID: 32720682
- PMCID: PMC7475651
- DOI: 10.1042/EBC20190074
CENP-A nucleosome-a chromatin-embedded pedestal for the centromere: lessons learned from structural biology
Abstract
The centromere is a chromosome locus that directs equal segregation of chromosomes during cell division. A nucleosome containing the histone H3 variant CENP-A epigenetically defines the centromere. Here, we summarize findings from recent structural biology studies, including several CryoEM structures, that contributed to elucidate specific features of the CENP-A nucleosome and molecular determinants of its interactions with CENP-C and CENP-N, the only two centromere proteins that directly bind to it. Based on those findings, we propose a role of the CENP-A nucleosome in the organization of centromeric chromatin beyond binding centromeric proteins.
Keywords: CENP-A; centromere; chromatin.
© 2020 The Author(s).
Conflict of interest statement
The authors declare that there are no competing interests associated with the manuscript.
Figures
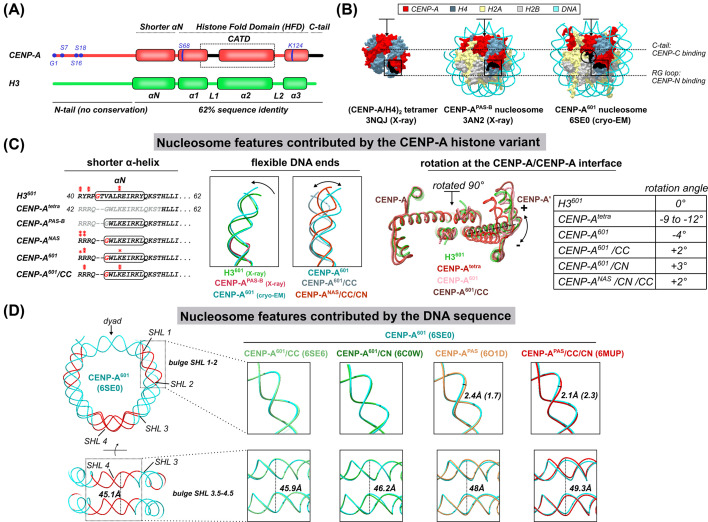
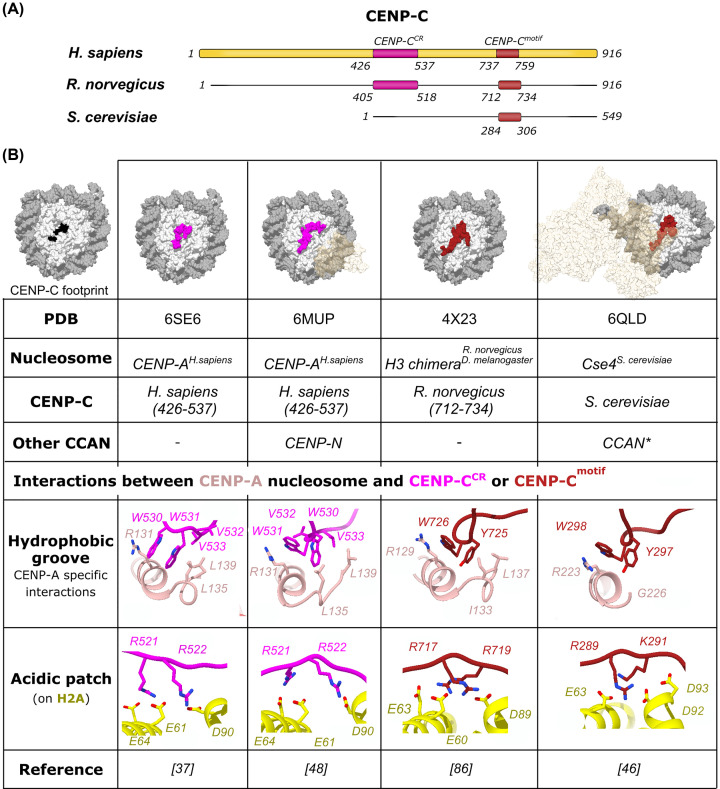
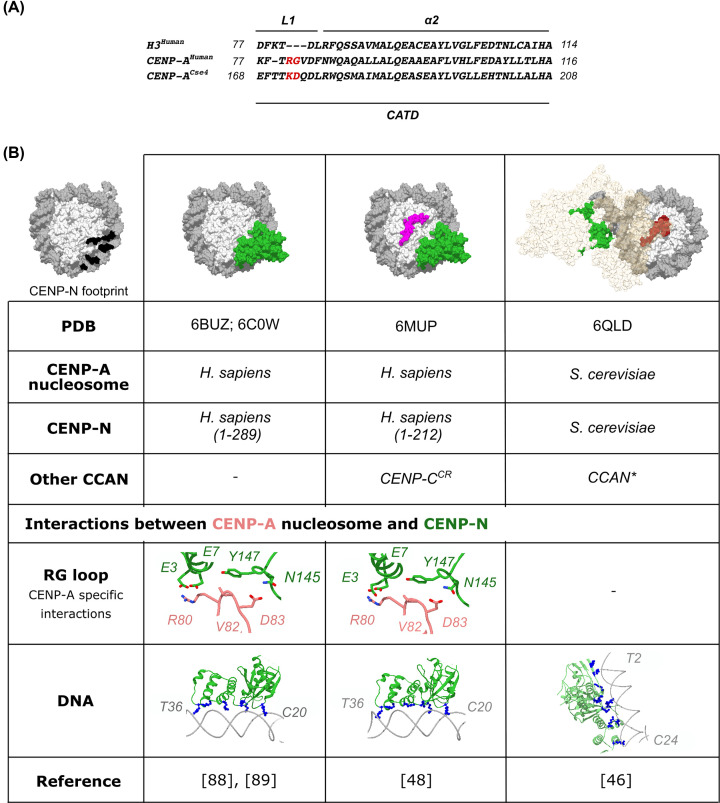
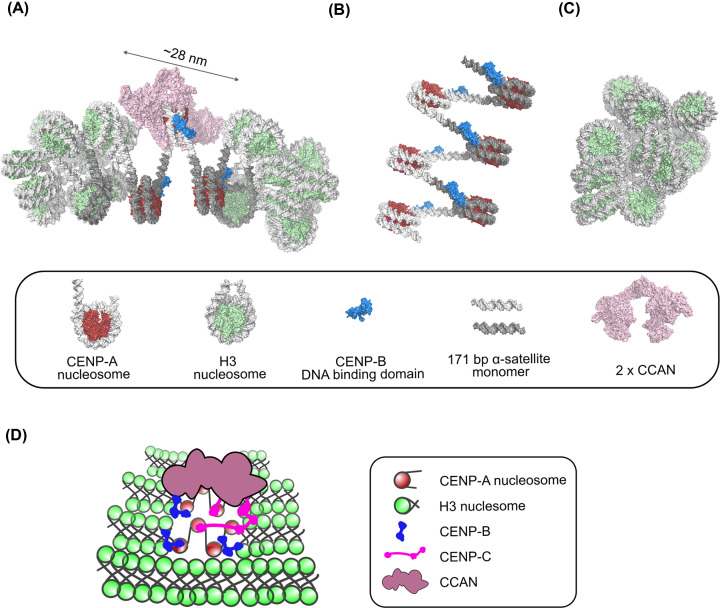
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources