

Enhanced MAPK1 Function Causes a Neurodevelopmental Disorder within the RASopathy Clinical Spectrum
- PMID: 32721402
- PMCID: PMC7477014
- DOI: 10.1016/j.ajhg.2020.06.018
Enhanced MAPK1 Function Causes a Neurodevelopmental Disorder within the RASopathy Clinical Spectrum
Abstract
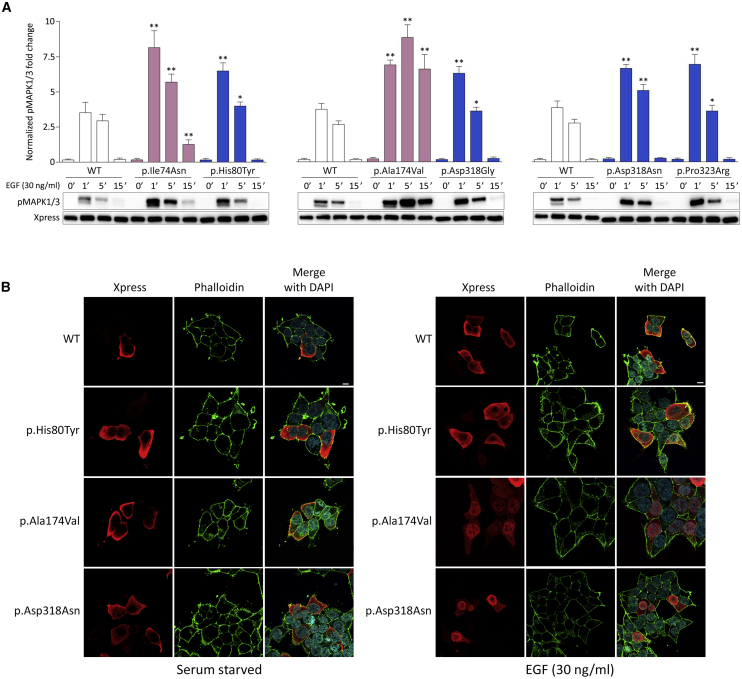
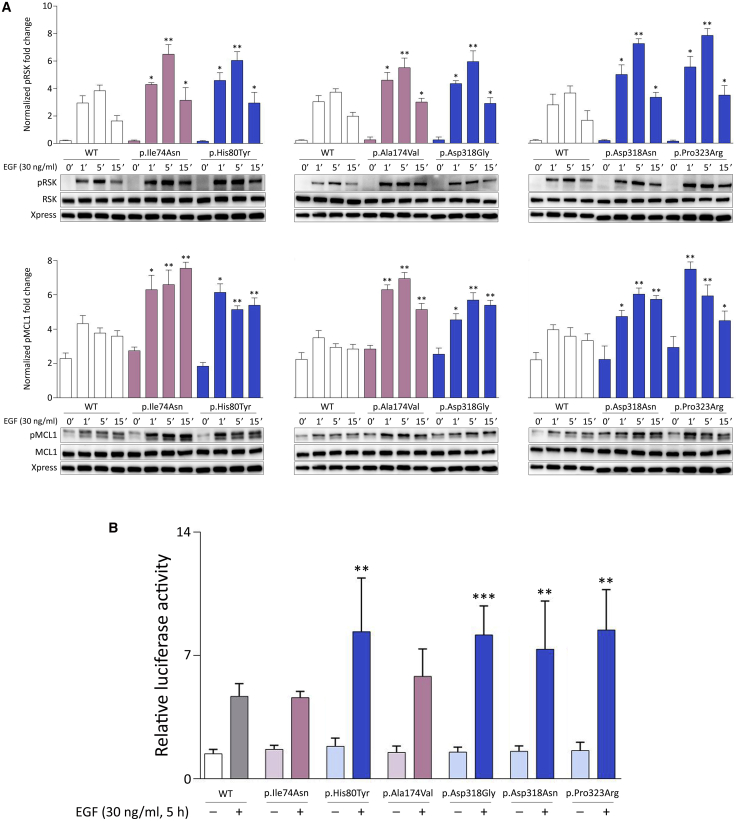
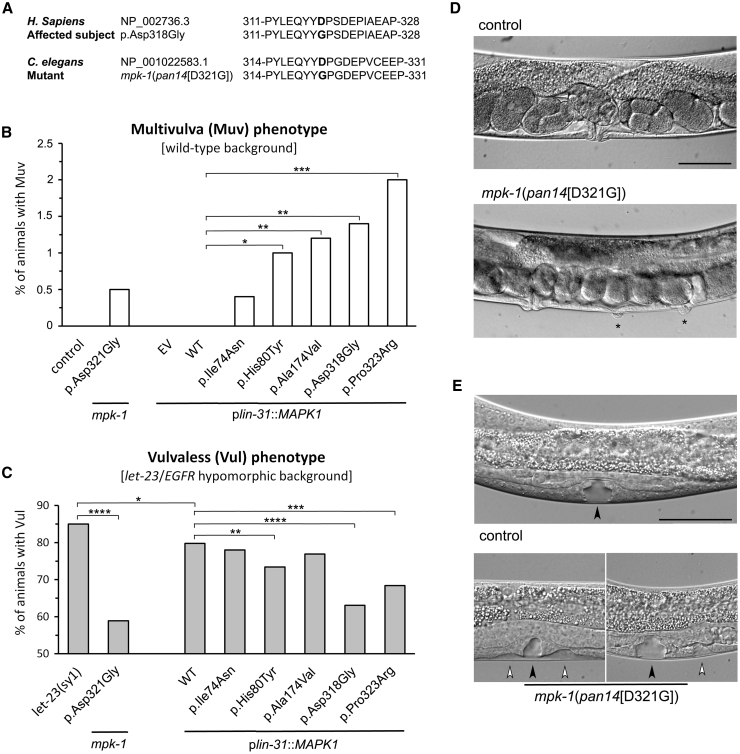
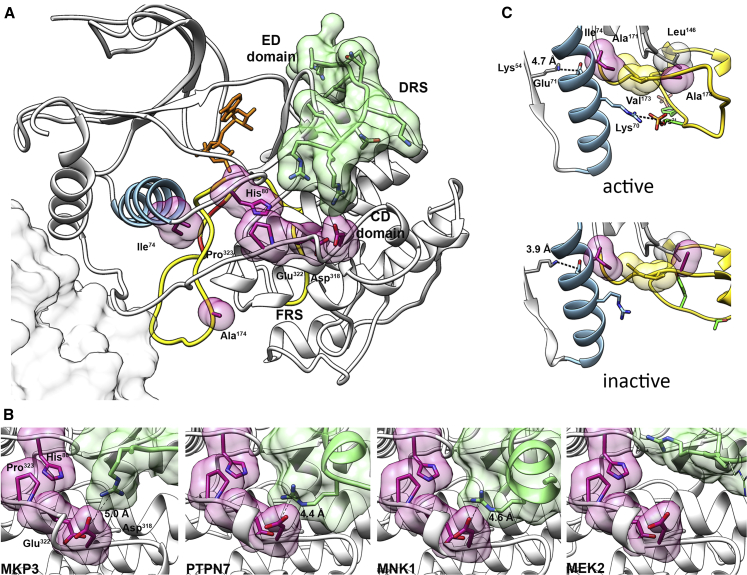
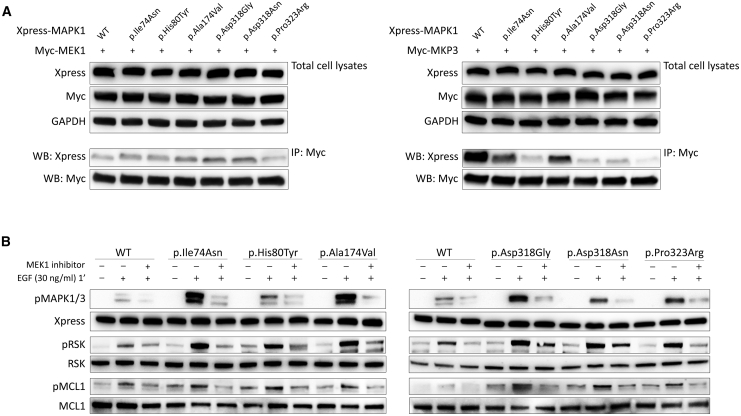
Signal transduction through the RAF-MEK-ERK pathway, the first described mitogen-associated protein kinase (MAPK) cascade, mediates multiple cellular processes and participates in early and late developmental programs. Aberrant signaling through this cascade contributes to oncogenesis and underlies the RASopathies, a family of cancer-prone disorders. Here, we report that de novo missense variants in MAPK1, encoding the mitogen-activated protein kinase 1 (i.e., extracellular signal-regulated protein kinase 2, ERK2), cause a neurodevelopmental disease within the RASopathy phenotypic spectrum, reminiscent of Noonan syndrome in some subjects. Pathogenic variants promote increased phosphorylation of the kinase, which enhances translocation to the nucleus and boosts MAPK signaling in vitro and in vivo. Two variant classes are identified, one of which directly disrupts binding to MKP3, a dual-specificity protein phosphatase negatively regulating ERK function. Importantly, signal dysregulation driven by pathogenic MAPK1 variants is stimulus reliant and retains dependence on MEK activity. Our data support a model in which the identified pathogenic variants operate with counteracting effects on MAPK1 function by differentially impacting the ability of the kinase to interact with regulators and substrates, which likely explains the minor role of these variants as driver events contributing to oncogenesis. After nearly 20 years from the discovery of the first gene implicated in Noonan syndrome, PTPN11, the last tier of the MAPK cascade joins the group of genes mutated in RASopathies.
Keywords: C. elegans; ERK2; MAPK cascade; MKP3; Noonan syndrome; RAS signaling; RASopathies; RSK; exome sequencing; intracellular signaling.
Copyright © 2020 American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
I.M.W. and M.M.M. are employees of GeneDx. S.Á. and M.M.-G. are employees of NimGenetics. All the other authors declare no competing interests.
Figures

References
-
- Yoon S., Seger R. The extracellular signal-regulated kinase: multiple substrates regulate diverse cellular functions. Growth Factors. 2006;24:21–44. - PubMed
-
- Kolch W. Coordinating ERK/MAPK signalling through scaffolds and inhibitors. Nat. Rev. Mol. Cell Biol. 2005;6:827–837. - PubMed
-
- Samatar A.A., Poulikakos P.I. Targeting RAS-ERK signalling in cancer: promises and challenges. Nat. Rev. Drug Discov. 2014;13:928–942. - PubMed
-
- Krens S.F., Spaink H.P., Snaar-Jagalska B.E. Functions of the MAPK family in vertebrate-development. FEBS Lett. 2006;580:4984–4990. - PubMed
-
- Hayashi S., Ogura Y. ERK signaling dynamics in the morphogenesis and homeostasis of Drosophila. Curr. Opin. Genet. Dev. 2020;63:9–15. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
