

Fatty Acid Allosteric Regulation of C-H Activation in Plant and Animal Lipoxygenases
- PMID: 32722330
- PMCID: PMC7436259
- DOI: 10.3390/molecules25153374
Fatty Acid Allosteric Regulation of C-H Activation in Plant and Animal Lipoxygenases
Abstract
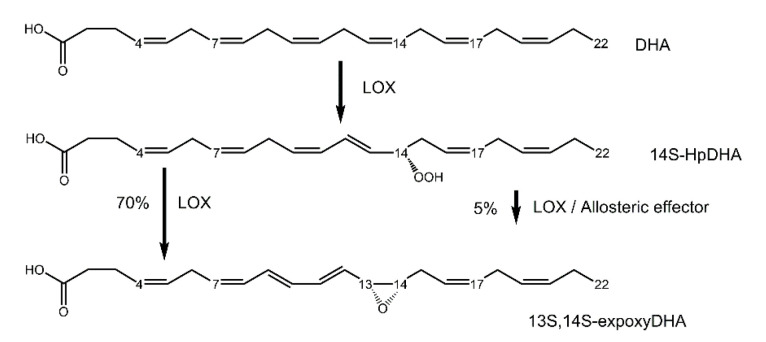
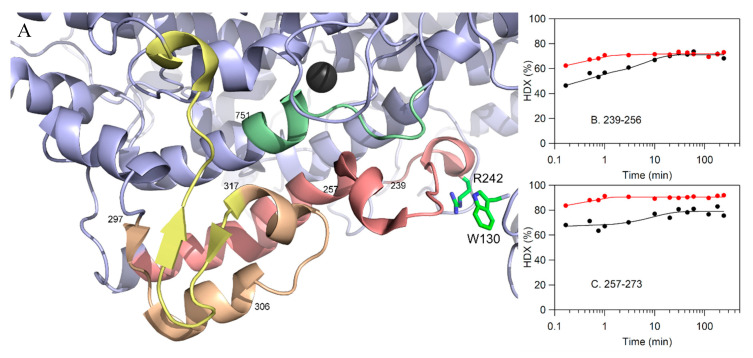
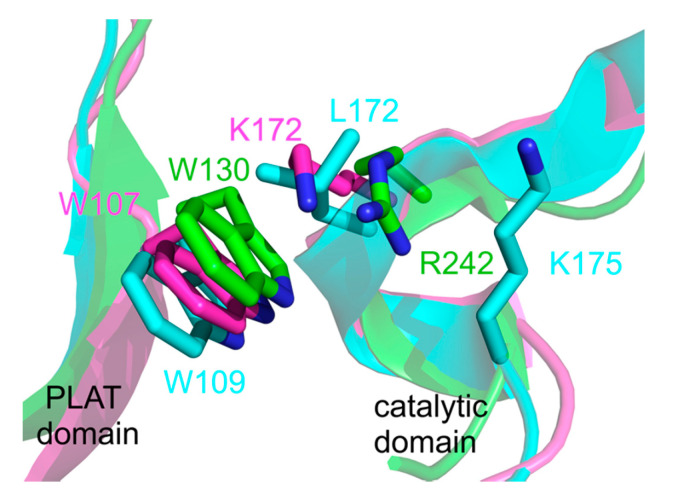
Lipoxygenases (LOXs) catalyze the (per) oxidation of fatty acids that serve as important mediators for cell signaling and inflammation. These reactions are initiated by a C-H activation step that is allosterically regulated in plant and animal enzymes. LOXs from higher eukaryotes are equipped with an N-terminal PLAT (Polycystin-1, Lipoxygenase, Alpha-Toxin) domain that has been implicated to bind to small molecule allosteric effectors, which in turn modulate substrate specificity and the rate-limiting steps of catalysis. Herein, the kinetic and structural evidence that describes the allosteric regulation of plant and animal lipoxygenase chemistry by fatty acids and their derivatives are summarized.
Keywords: C-H activation; hydrogen tunneling; kinetic isotope effects; protein allostery; substrate selectivity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures










References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
