

A Cytoplasmic Heme Sensor Illuminates the Impacts of Mitochondrial and Vacuolar Functions and Oxidative Stress on Heme-Iron Homeostasis in Cryptococcus neoformans
- PMID: 32723917
- PMCID: PMC7387795
- DOI: 10.1128/mBio.00986-20
A Cytoplasmic Heme Sensor Illuminates the Impacts of Mitochondrial and Vacuolar Functions and Oxidative Stress on Heme-Iron Homeostasis in Cryptococcus neoformans
Abstract
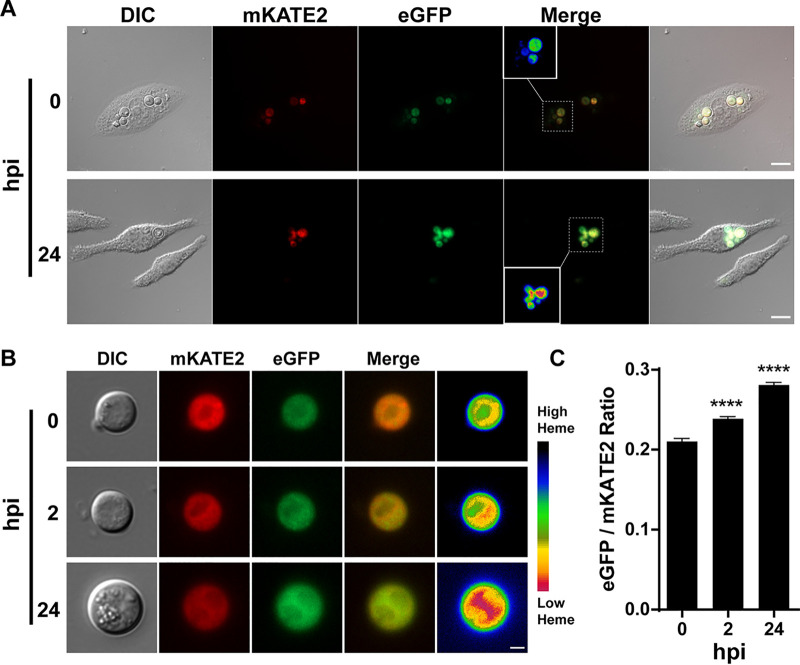
Pathogens must compete with hosts to acquire sufficient iron for proliferation during pathogenesis. The pathogenic fungus Cryptococcus neoformans is capable of acquiring iron from heme, the most abundant source in vertebrate hosts, although the mechanisms of heme sensing and acquisition are not entirely understood. In this study, we adopted a chromosomally encoded heme sensor developed for Saccharomyces cerevisiae to examine cytosolic heme levels in C. neoformans using fluorescence microscopy, fluorimetry, and flow cytometry. We validated the responsiveness of the sensor upon treatment with exogenous hemin, during proliferation in macrophages, and in strains defective for endocytosis. We then used the sensor to show that vacuolar and mitochondrial dysregulation and oxidative stress reduced the labile heme pool in the cytosol. Importantly, the sensor provided a tool to further demonstrate that the drugs artemisinin and metformin have heme-related activities and the potential to be repurposed for antifungal therapy. Overall, this study provides insights into heme sensing by C. neoformans and establishes a powerful tool to further investigate mechanisms of heme-iron acquisition in the context of fungal pathogenesis.IMPORTANCE Invasive fungal diseases are increasing in frequency, and new drug targets and antifungal drugs are needed to bolster therapy. The mechanisms by which pathogens obtain critical nutrients such as iron from heme during host colonization represent a promising target for therapy. In this study, we employed a fluorescent heme sensor to investigate heme homeostasis in Cryptococcus neoformans We demonstrated that endocytosis is a key aspect of heme acquisition and that vacuolar and mitochondrial functions are important in regulating the pool of available heme in cells. Stress generated by oxidative conditions impacts the heme pool, as do the drugs artemisinin and metformin; these drugs have heme-related activities and are in clinical use for malaria and diabetes, respectively. Overall, our study provides insights into mechanisms of fungal heme acquisition and demonstrates the utility of the heme sensor for drug characterization in support of new therapies for fungal diseases.
Keywords: artemisinin; biosensors; heme sensor; heme-related drugs; macrophage; metformin; mitochondria; reactive oxygen species; vacuole.
Copyright © 2020 Bairwa et al.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
