

Keratinocytes contribute to normal cold and heat sensation
- PMID: 32729832
- PMCID: PMC7402674
- DOI: 10.7554/eLife.58625
Keratinocytes contribute to normal cold and heat sensation
Abstract
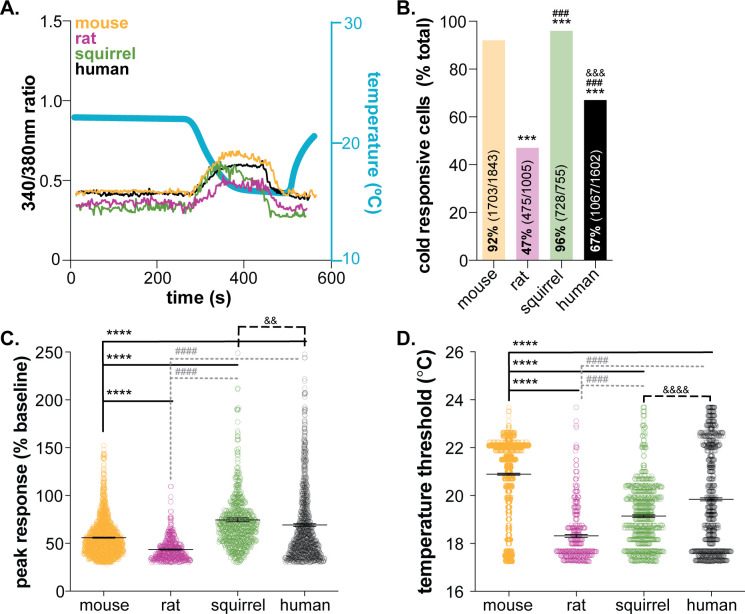
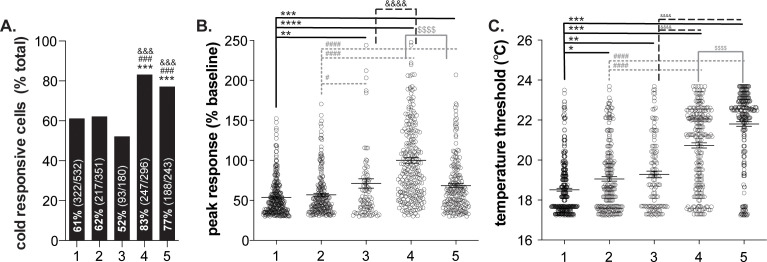
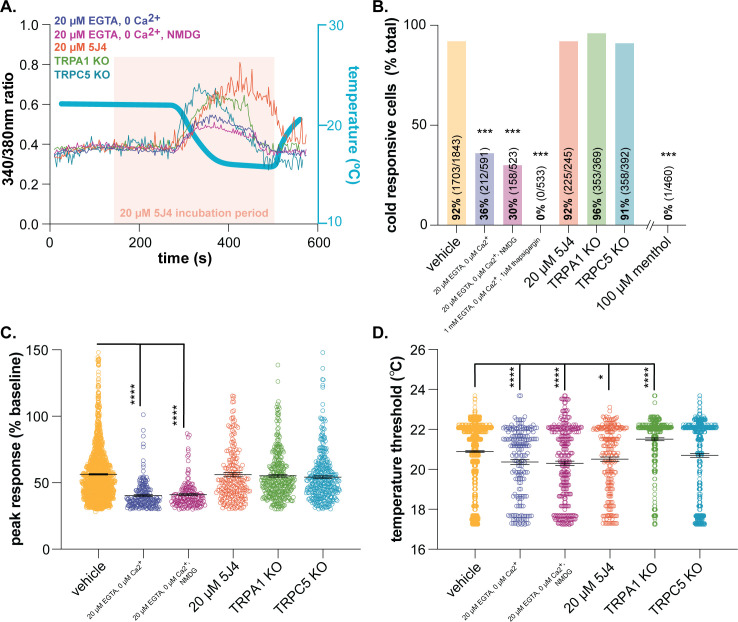
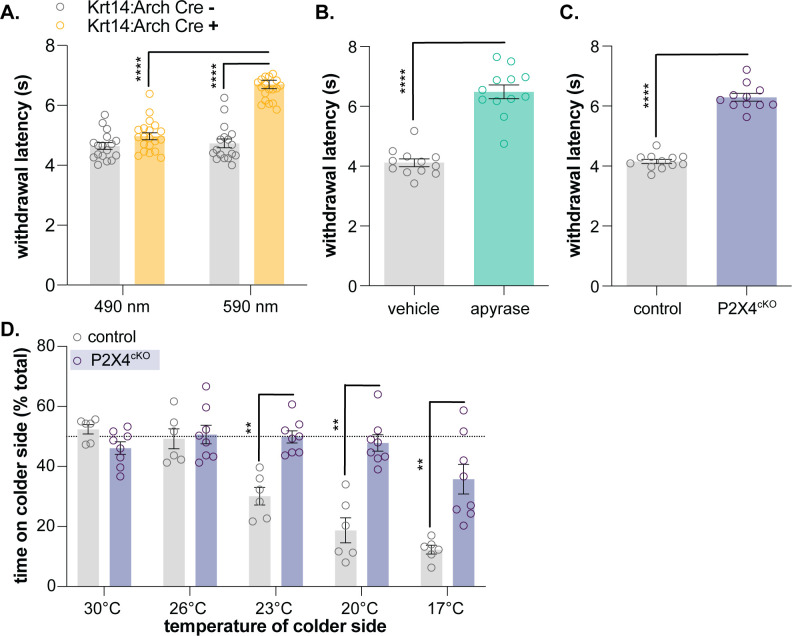
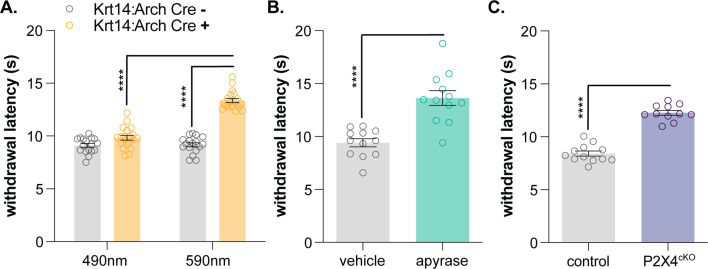
Keratinocytes are the most abundant cell type in the epidermis, the most superficial layer of skin. Historically, epidermal-innervating sensory neurons were thought to be the exclusive detectors and transmitters of environmental stimuli. However, recent work from our lab (Moehring et al., 2018) and others (Baumbauer et al., 2015) has demonstrated that keratinocytes are also critical for normal mechanotransduction and mechanically-evoked behavioral responses in mice. Here, we asked whether keratinocyte activity is also required for normal cold and heat sensation. Using calcium imaging, we determined that keratinocyte cold activity is conserved across mammalian species and requires the release of intracellular calcium through one or more unknown cold-sensitive proteins. Both epidermal cell optogenetic inhibition and interruption of ATP-P2X4 signaling reduced reflexive behavioral responses to cold and heat stimuli. Based on these data and our previous findings, keratinocyte purinergic signaling is a modality-conserved amplification system that is required for normal somatosensation in vivo.
Keywords: ATP; P2X4; cold; heat; human; keratinocyte; mouse; neuroscience; rat.
© 2020, Sadler et al.
Conflict of interest statement
KS, FM, CS No competing interests declared
Figures
References
-
- Bidaux G, Borowiec AS, Gordienko D, Beck B, Shapovalov GG, Lemonnier L, Flourakis M, Vandenberghe M, Slomianny C, Dewailly E, Delcourt P, Desruelles E, Ritaine A, Polakowska R, Lesage J, Chami M, Skryma R, Prevarskaya N. Epidermal TRPM8 channel isoform controls the balance between keratinocyte proliferation and differentiation in a cold-dependent manner. PNAS. 2015;112:E3345–E3354. doi: 10.1073/pnas.1423357112. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
