

DCAF1 regulates Treg senescence via the ROS axis during immunological aging
- PMID: 32730228
- PMCID: PMC7598062
- DOI: 10.1172/JCI136466
DCAF1 regulates Treg senescence via the ROS axis during immunological aging
Abstract
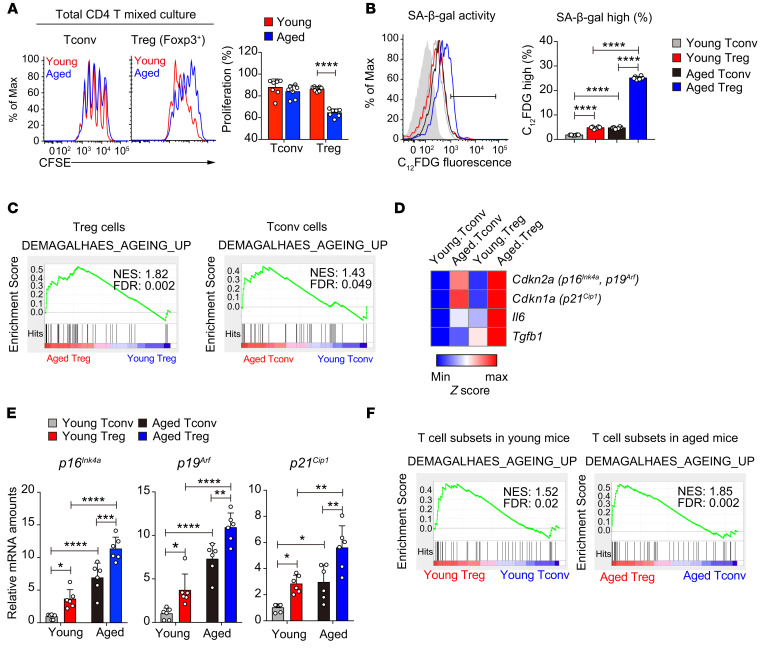
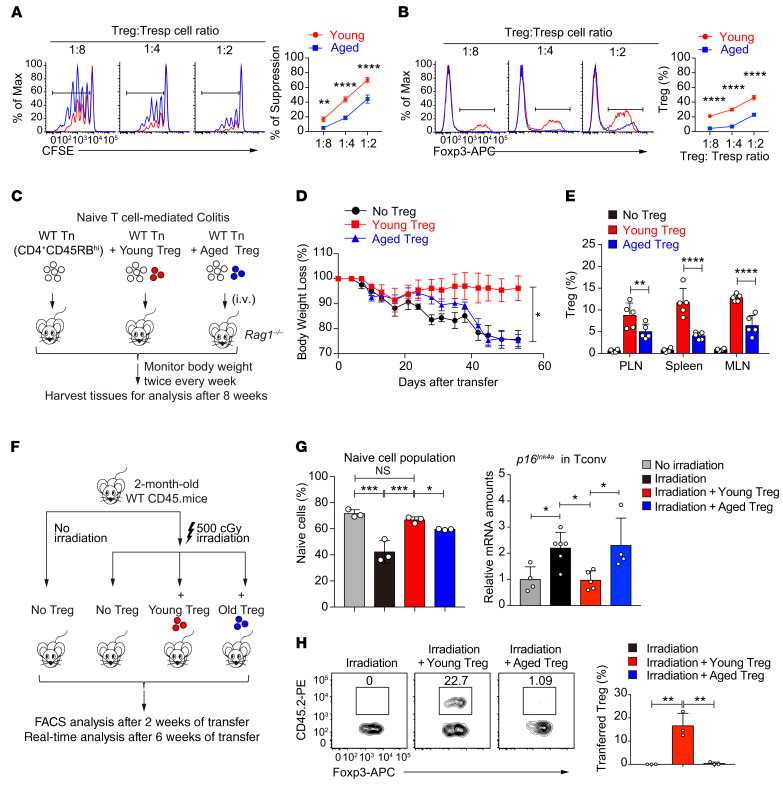
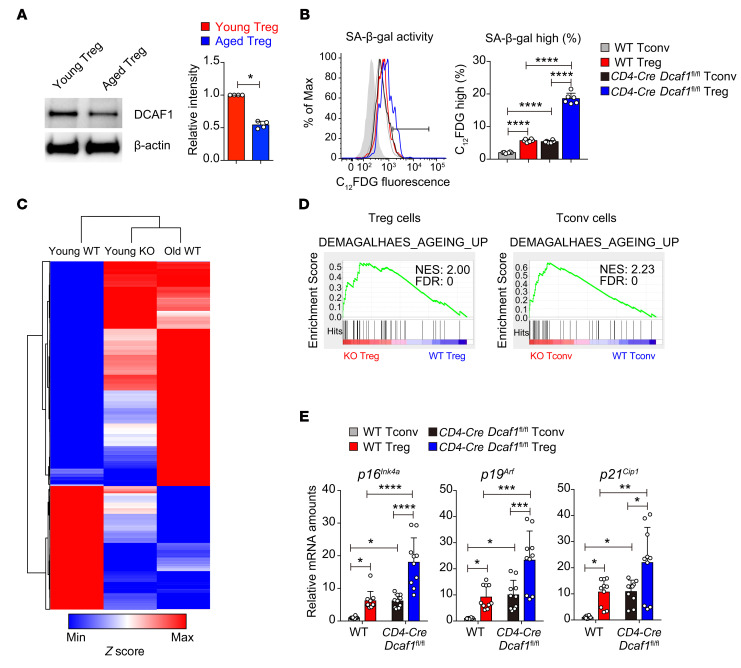
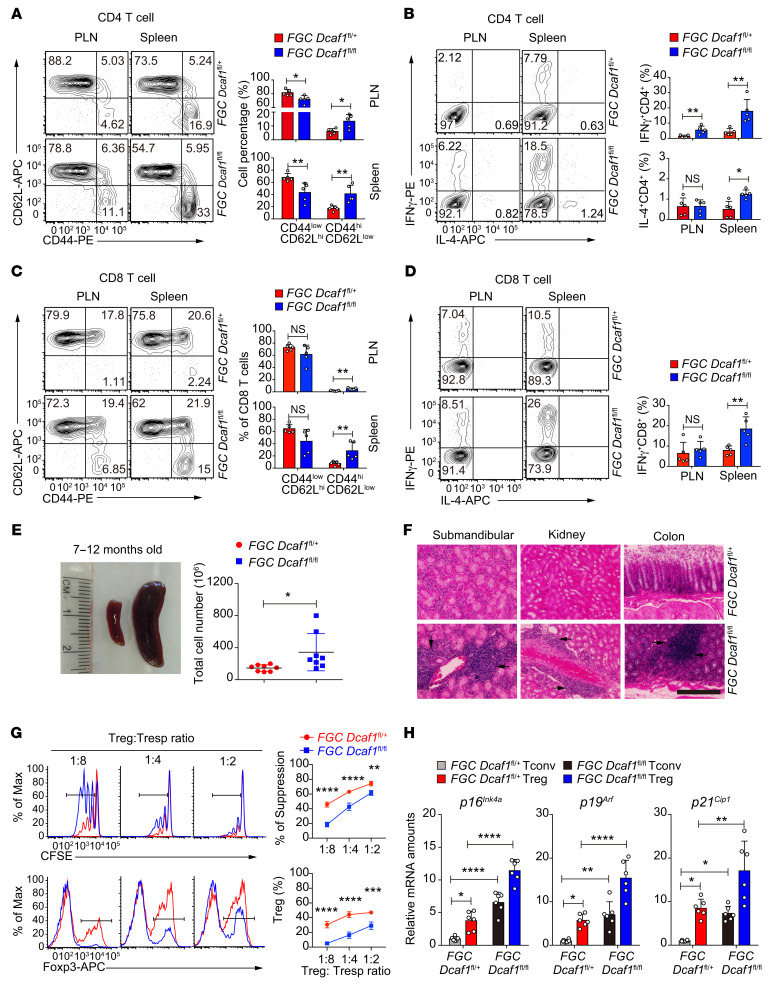
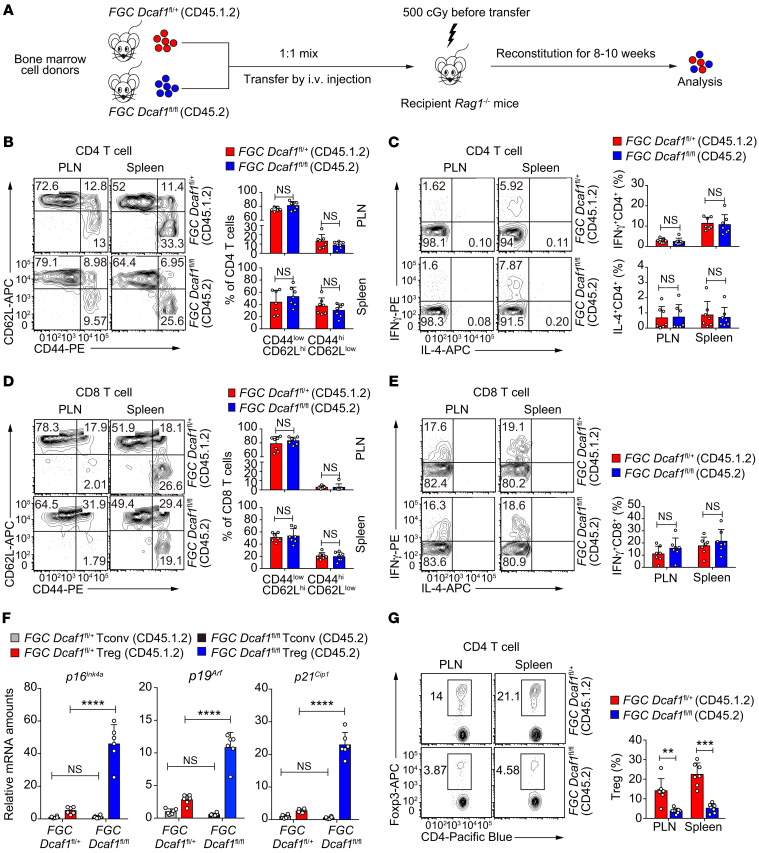
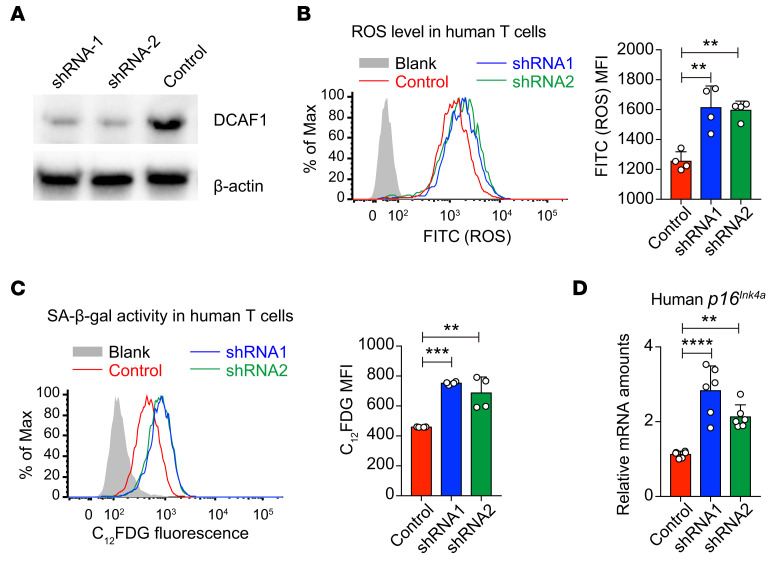
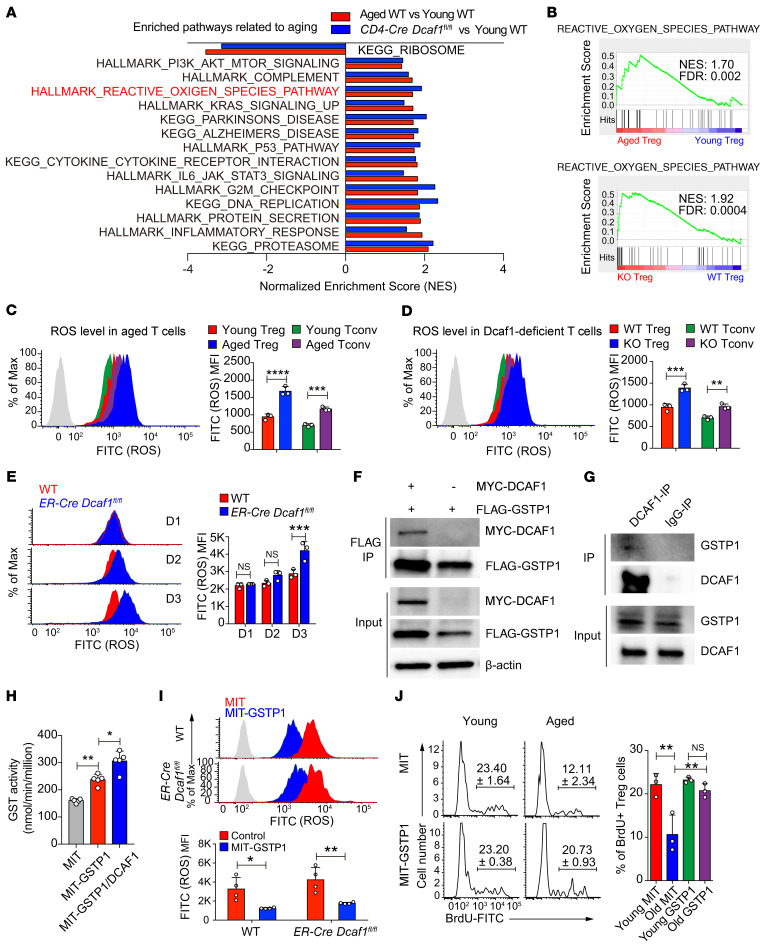
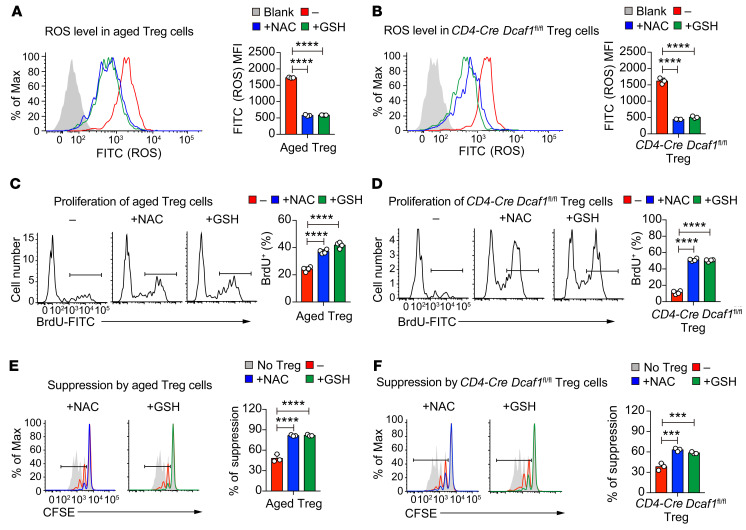
As a hallmark of immunological aging, low-grade, chronic inflammation with accumulation of effector memory T cells contributes to increased susceptibility to many aging-related diseases. While the proinflammatory state of aged T cells indicates a dysregulation of immune homeostasis, whether and how aging drives regulatory T cell (Treg) aging and alters Treg function are not fully understood owing to a lack of specific aging markers. Here, by a combination of cellular, molecular, and bioinformatic approaches, we discovered that Tregs senesce more severely than conventional T (Tconv) cells during aging. We found that Tregs from aged mice were less efficient than young Tregs in suppressing Tconv cell function in an inflammatory bowel disease model and in preventing Tconv cell aging in an irradiation-induced aging model. Furthermore, we revealed that DDB1- and CUL4-associated factor 1 (DCAF1) was downregulated in aged Tregs and was critical to restrain Treg aging via reactive oxygen species (ROS) regulated by glutathione-S-transferase P (GSTP1). Importantly, interfering with GSTP1 and ROS pathways reinvigorated the proliferation and function of aged Tregs. Therefore, our studies uncover an important role of the DCAF1/GSTP1/ROS axis in Treg senescence, which leads to uncontrolled inflammation and immunological aging.
Keywords: Aging; Cellular senescence; Immunology; Inflammatory bowel disease; T cells.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
