

Introns mediate post-transcriptional enhancement of nuclear gene expression in the green microalga Chlamydomonas reinhardtii
- PMID: 32730252
- PMCID: PMC7419008
- DOI: 10.1371/journal.pgen.1008944
Introns mediate post-transcriptional enhancement of nuclear gene expression in the green microalga Chlamydomonas reinhardtii
Abstract
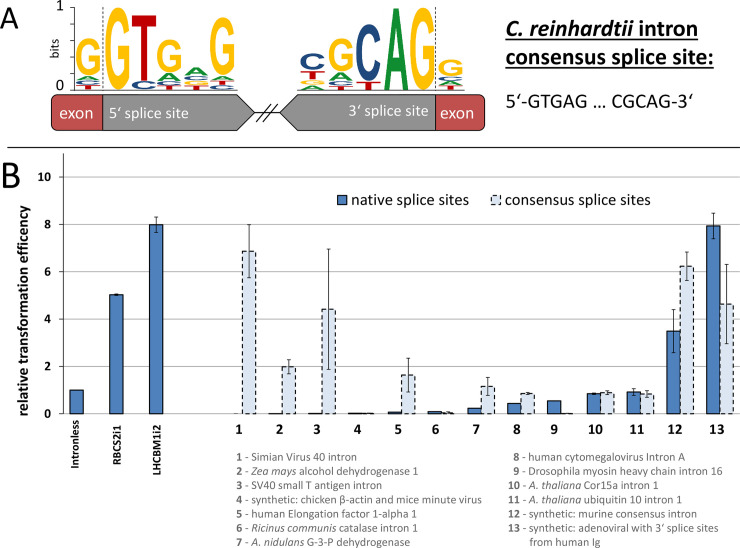
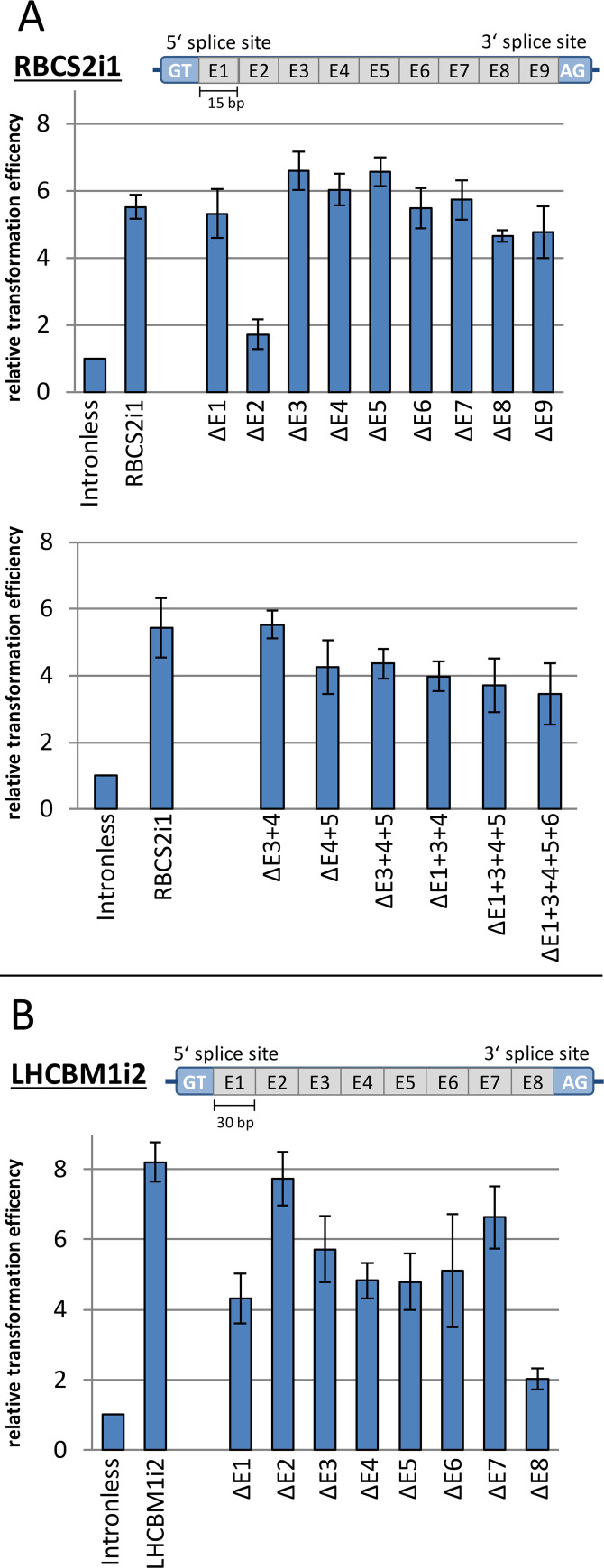
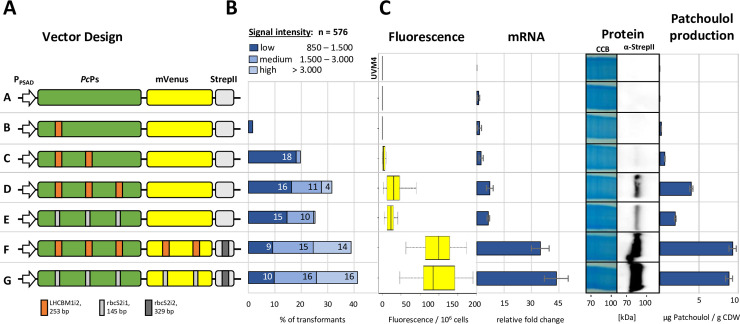
Efficient nuclear transgene expression in the green microalga Chlamydomonas reinhardtii is generally hindered by low transcription rates. Introns can increase transcript abundance by a process called Intron-Mediated Enhancement (IME) in this alga and has been broadly observed in other eukaryotes. However, the mechanisms of IME in microalgae are poorly understood. Here, we identified 33 native introns from highly expressed genes in C. reinhardtii selected from transcriptome studies as well as 13 non-native introns. We investigated their IME capacities and probed the mechanism of action by modification of splice sites, internal sequence motifs, and position within transgenes. Several introns were found to elicit strong IME and found to be broadly applicable in different expression constructs. We determined that IME in C. reinhardtii exclusively occurs from introns within transcribed ORFs regardless of the promoter and is not induced by traditional enhancers of transcription. Our results elucidate some mechanistic details of IME in C. reinhardtii, which are similar to those observed in higher plants yet underly distinctly different induction processes. Our findings narrow the focus of targets responsible for algal IME and provides evidence that introns are underestimated regulators of C. reinhardtii nuclear gene expression.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
