

Sex specific effects of "junk-food" diet on calcium permeable AMPA receptors and silent synapses in the nucleus accumbens core
- PMID: 32731252
- PMCID: PMC8027187
- DOI: 10.1038/s41386-020-0781-1
Sex specific effects of "junk-food" diet on calcium permeable AMPA receptors and silent synapses in the nucleus accumbens core
Abstract
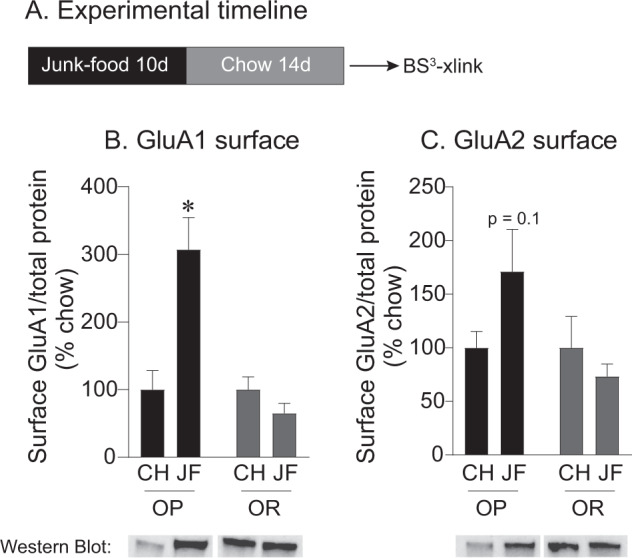
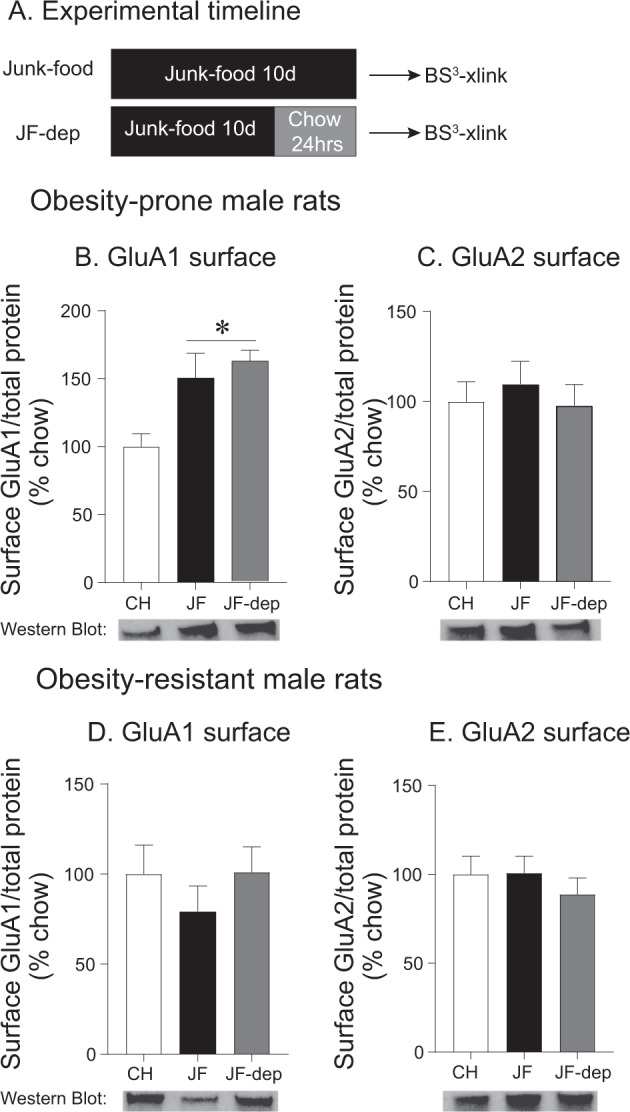
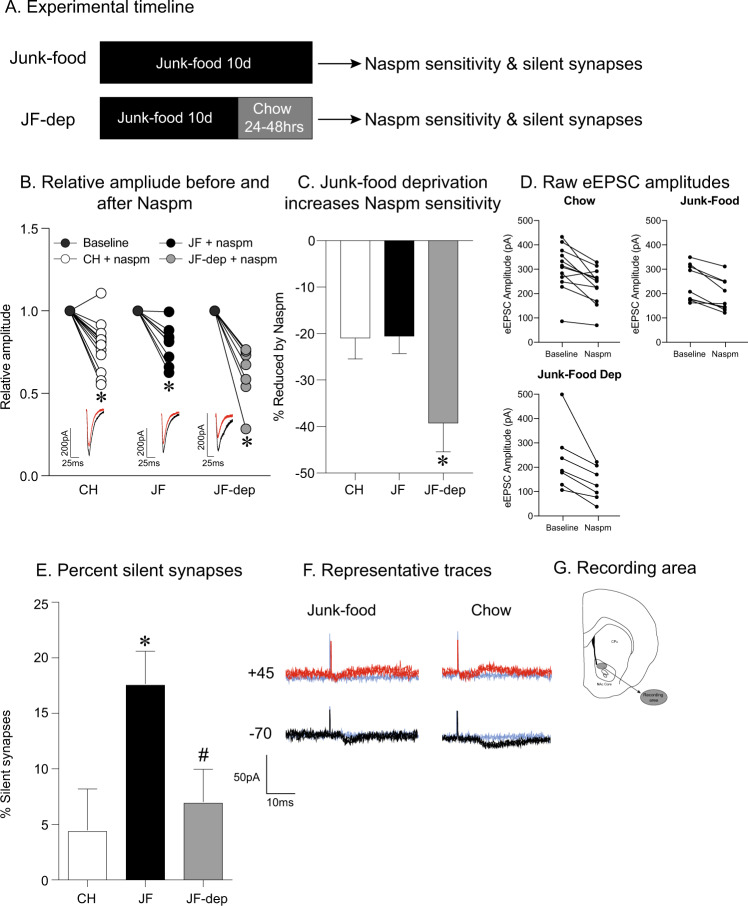
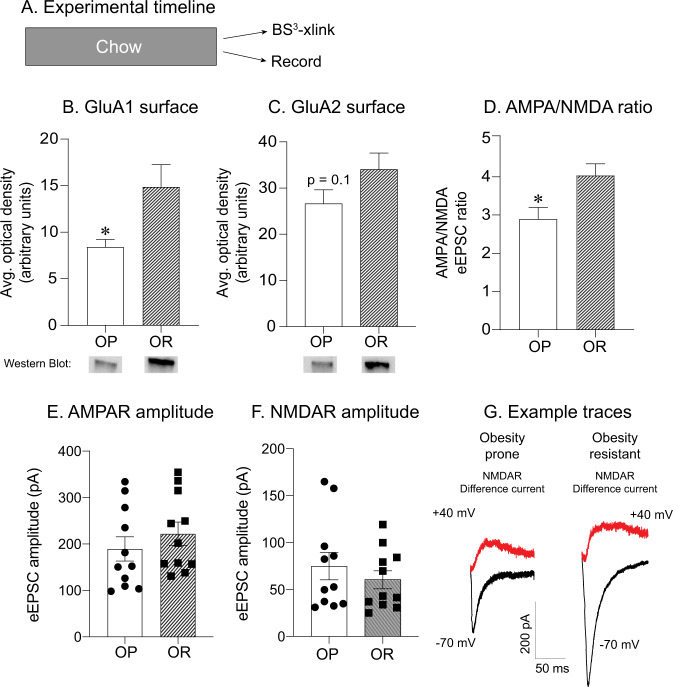
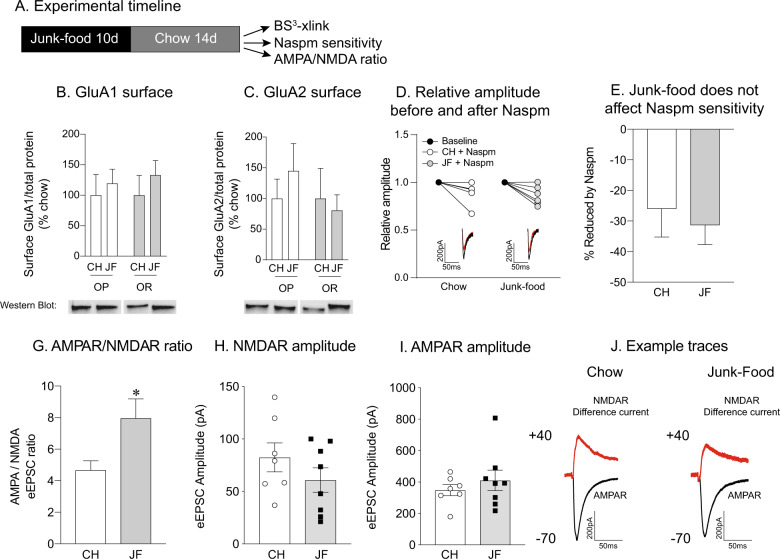
CP-AMPARs in the nucleus accumbens (NAc) mediate cue-triggered motivation for food and cocaine. In addition, increases in NAc CP-AMPAR expression and function can be induced by cocaine or sugary, fatty junk-foods. However, the precise nature of these alterations and the degree to which they rely on the same underlying mechanisms is not well understood. This has important implications for understanding adaptive vs. maladaptive plasticity that drives food- and drug-seeking behaviors. Furthermore, effects of junk-foods on glutamatergic plasticity in females are unknown. Here, we use a combination of protein biochemistry and whole-cell patch clamping to determine effects of diet manipulation on glutamatergic plasticity within the NAc of males and females. We found that junk-food consumption increases silent synapses and subsequently increases CP-AMPAR levels in males in the NAc of male rats. In addition, a brief period of junk-food deprivation is needed for the synaptic insertion of CP-AMPARs and the maturation of silent synapses in males. In contrast, junk-food did not induce AMPAR plasticity in females but may instead alter NMDAR-mediated transmission. Thus, these studies reveal sex differences in the effects of junk-food on NAc synaptic plasticity. In addition, they provide novel insights into how essential food rewards alter NAc function.
Figures
References
-
- Kelley AE, Baldo BA, Pratt WE, Will MJ. Corticostriatal-hypothalamic circuitry and food motivation: integration of energy, action and reward. Physiol Behav. 2005;86:773–95. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
