

Iron homeostasis and plant immune responses: Recent insights and translational implications
- PMID: 32732287
- PMCID: PMC7521657
- DOI: 10.1074/jbc.REV120.010856
Iron homeostasis and plant immune responses: Recent insights and translational implications
Abstract
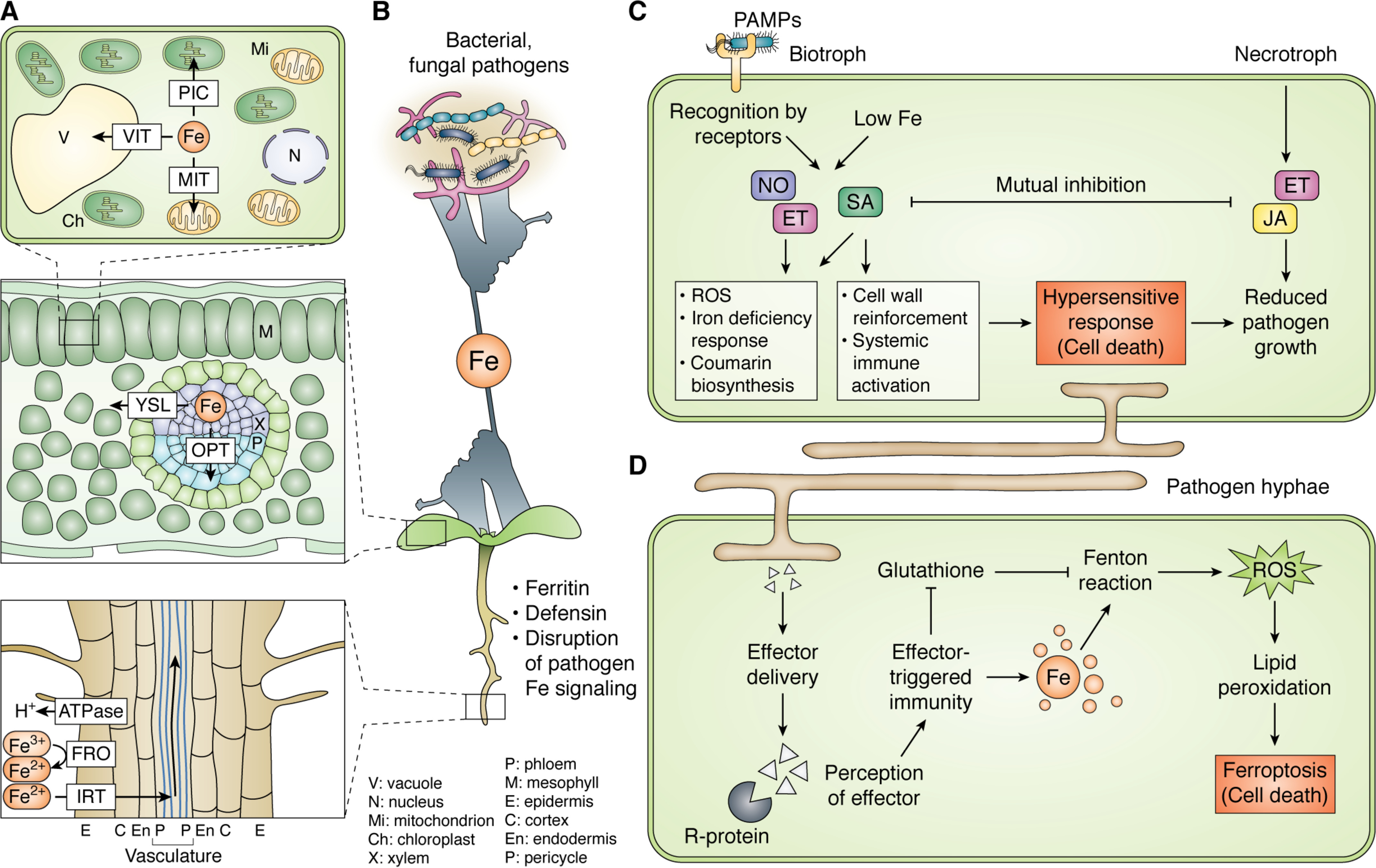
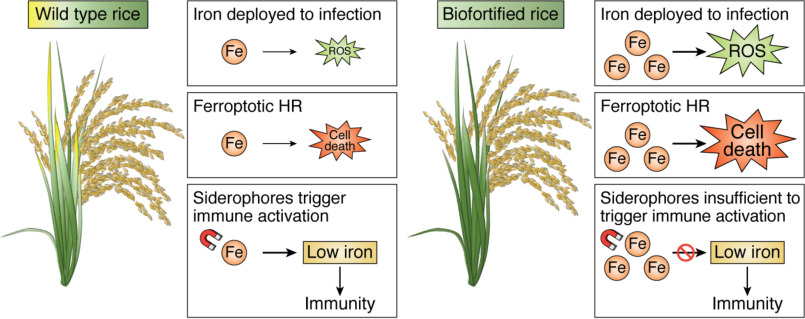
Iron metabolism and the plant immune system are both critical for plant vigor in natural ecosystems and for reliable agricultural productivity. Mechanistic studies of plant iron home-ostasis and plant immunity have traditionally been carried out in isolation from each other; however, our growing understanding of both processes has uncovered significant connections. For example, iron plays a critical role in the generation of reactive oxygen intermediates during immunity and has been recently implicated as a critical factor for immune-initiated cell death via ferroptosis. Moreover, plant iron stress triggers immune activation, suggesting that sensing of iron depletion is a mechanism by which plants recognize a pathogen threat. The iron deficiency response engages hormone signaling sectors that are also utilized for plant immune signaling, providing a probable explanation for iron-immunity cross-talk. Finally, interference with iron acquisition by pathogens might be a critical component of the immune response. Efforts to address the global burden of iron deficiency-related anemia have focused on classical breeding and transgenic approaches to develop crops biofortified for iron content. However, our improved mechanistic understanding of plant iron metabolism suggests that such alterations could promote or impede plant immunity, depending on the nature of the alteration and the virulence strategy of the pathogen. Effects of iron biofortification on disease resistance should be evaluated while developing plants for iron biofortification.
Keywords: Arabidopsis thaliana; host-pathogen interaction; iron metabolism; metal homeostasis; oxygen radicals; plant defense; plant hormone.
© 2020 Herlihy et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures
Similar articles
-
Immunity to plant pathogens and iron homeostasis.Plant Sci. 2015 Nov;240:90-7. doi: 10.1016/j.plantsci.2015.08.022. Epub 2015 Aug 29. Plant Sci. 2015. PMID: 26475190 Review.
-
Iron and Immunity.Annu Rev Phytopathol. 2017 Aug 4;55:355-375. doi: 10.1146/annurev-phyto-080516-035537. Epub 2017 Jun 9. Annu Rev Phytopathol. 2017. PMID: 28598721 Review.
-
Transcriptome landscape of a bacterial pathogen under plant immunity.Proc Natl Acad Sci U S A. 2018 Mar 27;115(13):E3055-E3064. doi: 10.1073/pnas.1800529115. Epub 2018 Mar 12. Proc Natl Acad Sci U S A. 2018. PMID: 29531038 Free PMC article.
-
New insights into the role of siderophores as triggers of plant immunity: what can we learn from animals?J Exp Bot. 2015 Jun;66(11):3001-10. doi: 10.1093/jxb/erv155. Epub 2015 May 1. J Exp Bot. 2015. PMID: 25934986 Review.
-
The role of iron in the immune response to bacterial infection.Immunol Res. 2011 May;50(1):1-9. doi: 10.1007/s12026-010-8199-1. Immunol Res. 2011. PMID: 21161695 Free PMC article. Review.
Cited by
-
Iron Availability in Tissue Microenvironment: The Key Role of Ferroportin.Int J Mol Sci. 2021 Mar 15;22(6):2986. doi: 10.3390/ijms22062986. Int J Mol Sci. 2021. PMID: 33804198 Free PMC article. Review.
-
Ferrous Sulfate-Mediated Control of Phytophthora capsici Pathogenesis and Its Impact on Pepper Plant.Plants (Basel). 2023 Dec 15;12(24):4168. doi: 10.3390/plants12244168. Plants (Basel). 2023. PMID: 38140495 Free PMC article.
-
Salicylic Acid-Mediated Disturbance Increases Bacterial Diversity in the Phyllosphere but Is Overcome by a Dominant Core Community.Front Microbiol. 2022 Feb 24;13:809940. doi: 10.3389/fmicb.2022.809940. eCollection 2022. Front Microbiol. 2022. PMID: 35283825 Free PMC article.
-
Iron Induces Resistance Against the Rice Blast Fungus Magnaporthe oryzae Through Potentiation of Immune Responses.Rice (N Y). 2022 Dec 25;15(1):68. doi: 10.1186/s12284-022-00609-w. Rice (N Y). 2022. PMID: 36566483 Free PMC article.
-
Genome analysis of a halophilic Virgibacillus halodenitrificans ASH15 revealed salt adaptation, plant growth promotion, and isoprenoid biosynthetic machinery.Front Microbiol. 2023 Sep 22;14:1229955. doi: 10.3389/fmicb.2023.1229955. eCollection 2023. Front Microbiol. 2023. PMID: 37808307 Free PMC article.
References
-
- Becana M., Moran J., and Iturbe-Ormaetxe I. (1998) Iron-dependent oxygen free radical generation in plants subjected to environmental stress: toxicity and antioxidant protection. Plant Soil 201, 137–147 10.1023/A:1004375732137 - DOI
-
- Pinto S. D S., Souza A. E. D., Oliva M. A., and Pereira E. G. (2016) Oxidative damage and photosynthetic impairment in tropical rice cultivars upon exposure to excess iron. Sci. Agric. 73, 217–226 10.1590/0103-9016-2015-0288 - DOI
-
- Tsai T.-M., and Huang H.-J. (2006) Effects of iron excess on cell viability and mitogen-activated protein kinase activation in rice roots. Physiol. Plant. 127, 583–592 10.1111/j.1399-3054.2006.00696.x - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
