

DNA capture by a CRISPR-Cas9-guided adenine base editor
- PMID: 32732424
- PMCID: PMC8598131
- DOI: 10.1126/science.abb1390
DNA capture by a CRISPR-Cas9-guided adenine base editor
Abstract
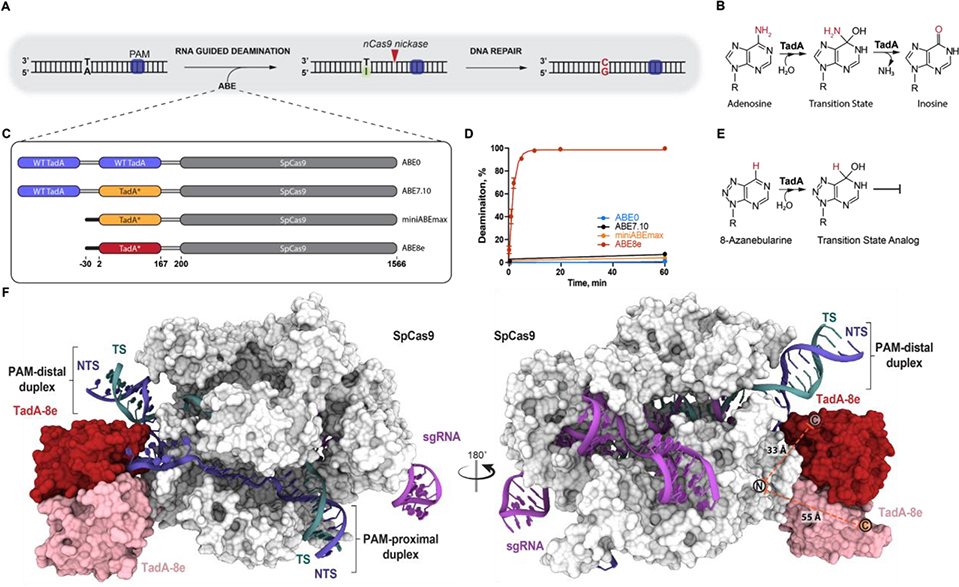
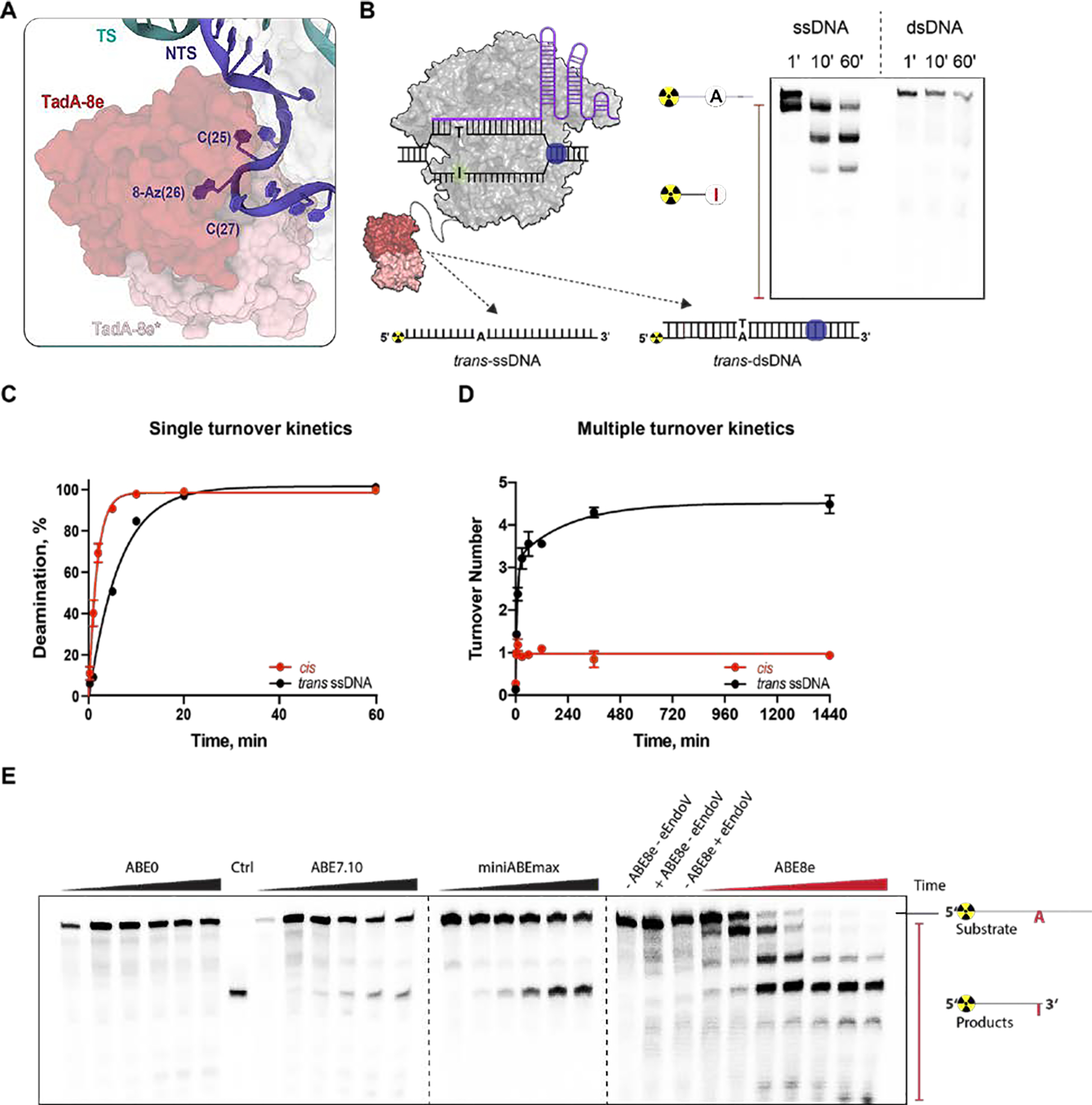
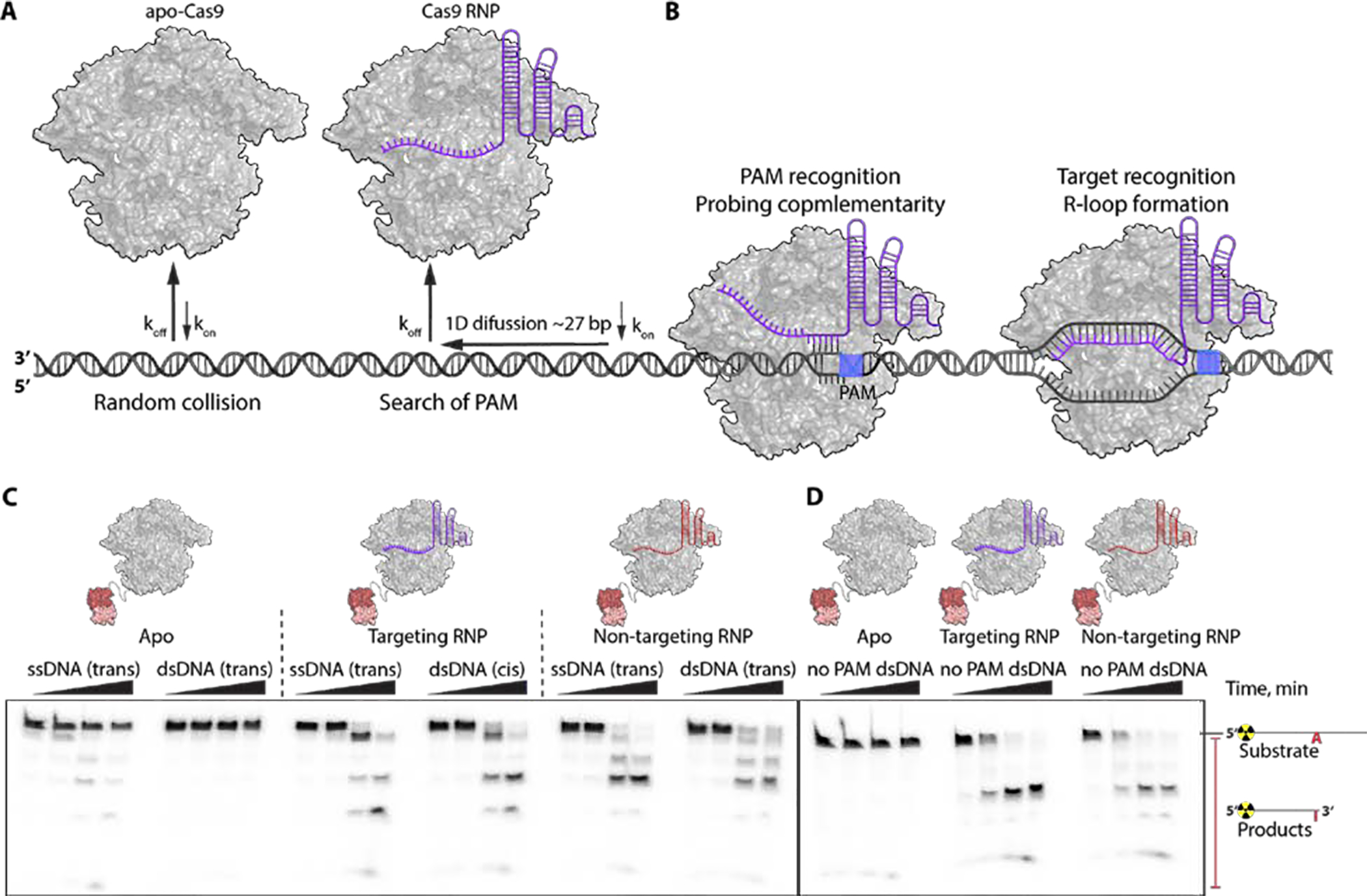
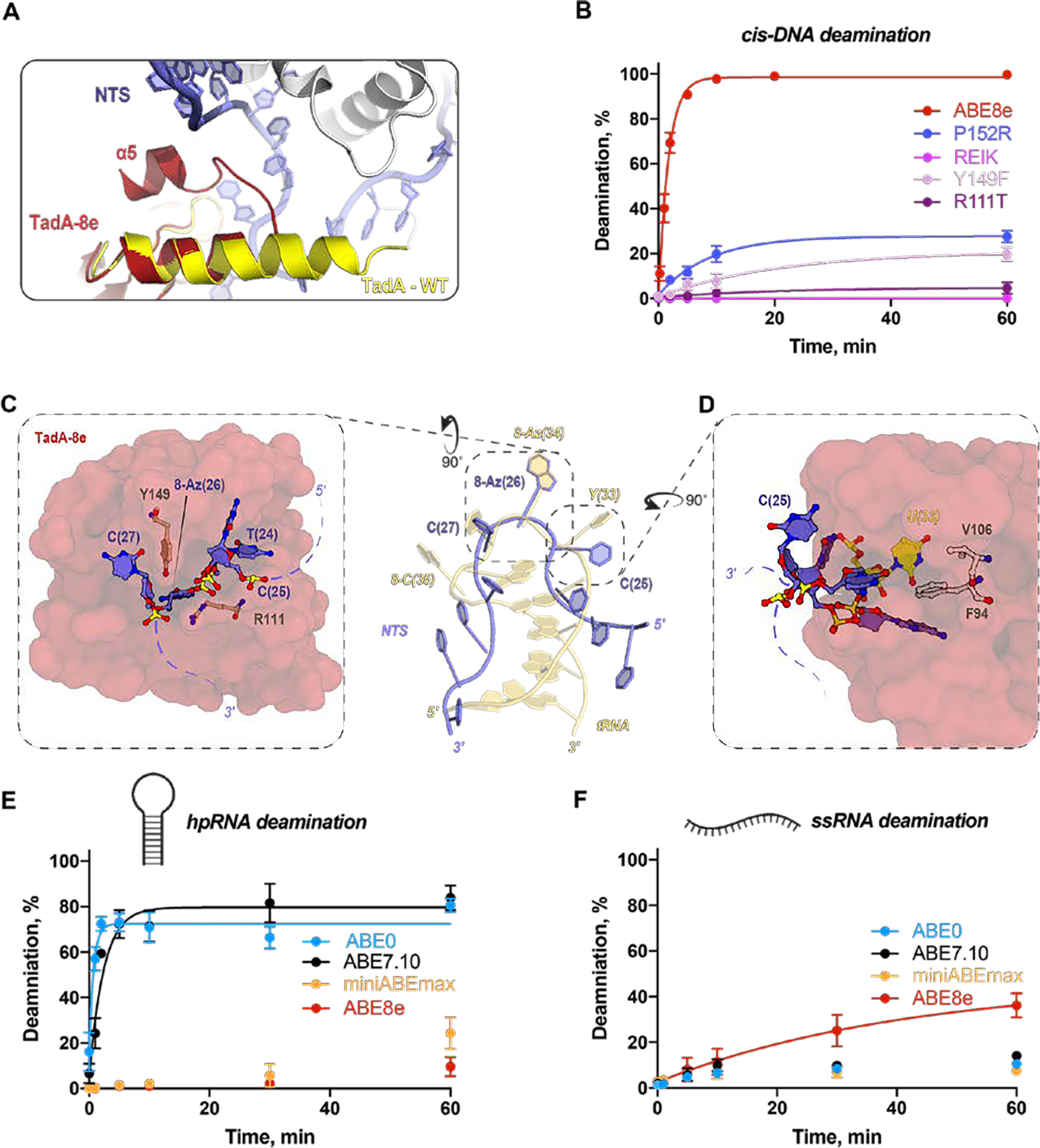
CRISPR-Cas-guided base editors convert A•T to G•C, or C•G to T•A, in cellular DNA for precision genome editing. To understand the molecular basis for DNA adenosine deamination by adenine base editors (ABEs), we determined a 3.2-angstrom resolution cryo-electron microscopy structure of ABE8e in a substrate-bound state in which the deaminase domain engages DNA exposed within the CRISPR-Cas9 R-loop complex. Kinetic and structural data suggest that ABE8e catalyzes DNA deamination up to ~1100-fold faster than earlier ABEs because of mutations that stabilize DNA substrates in a constrained, transfer RNA-like conformation. Furthermore, ABE8e's accelerated DNA deamination suggests a previously unobserved transient DNA melting that may occur during double-stranded DNA surveillance by CRISPR-Cas9. These results explain ABE8e-mediated base-editing outcomes and inform the future design of base editors.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
-
- Landrum MJ, Lee JM, Benson M, Brown G, Chao C, Chitipiralla S, Gu B, Hart J, Hoffman D, Hoover J, Jang W, Katz K, Ovetsky M, Riley G, Sethi A, Tully R, Villamarin-Salomon R, Rubinstein W, Maglott DR, ClinVar: public archive of interpretations of clinically relevant variants. Nucleic Acids Research. 44 (2016), pp. D862–D868. - PMC - PubMed
-
- Kim J, Malashkevich V, Roday S, Lisbin M, Schramm VL, Almo SC, Structural and Kinetic Characterization ofEscherichia coliTadA, the Wobble-Specific tRNA Deaminase†. Biochemistry. 45 (2006), pp. 6407–6416. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
