

In Vivo Selection for Formate Dehydrogenases with High Efficiency and Specificity toward NADP
- PMID: 32733773
- PMCID: PMC7384739
- DOI: 10.1021/acscatal.0c01487
In Vivo Selection for Formate Dehydrogenases with High Efficiency and Specificity toward NADP
Abstract
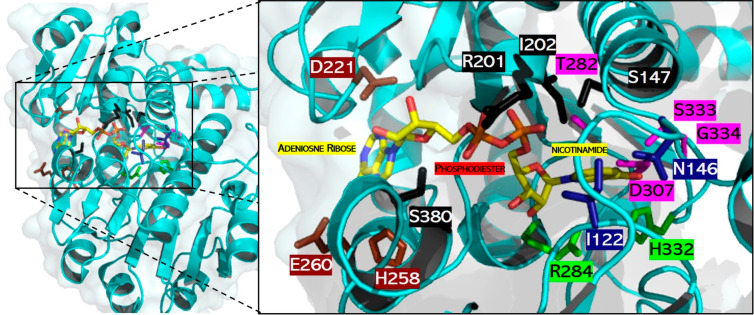
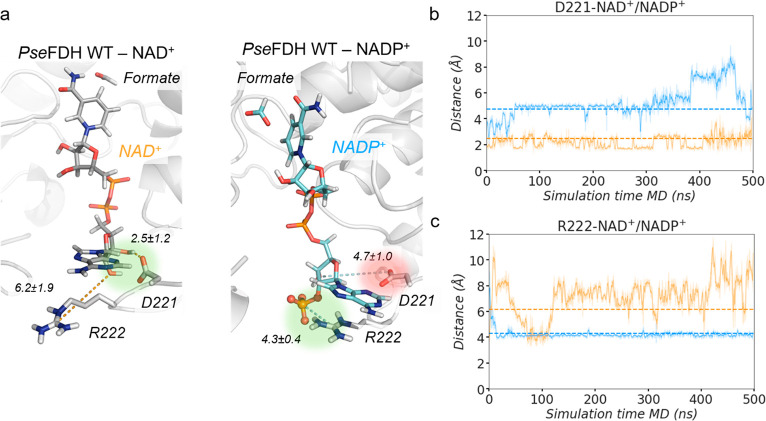
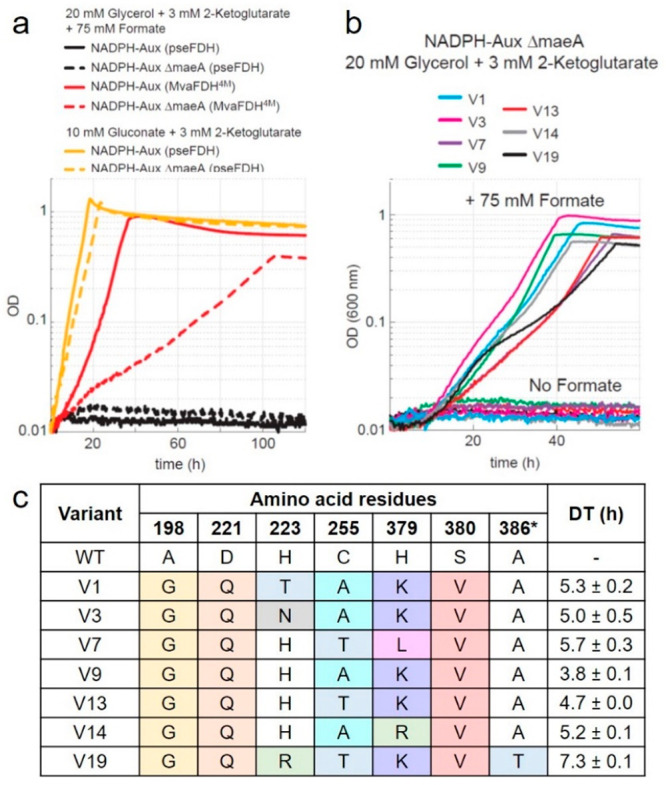
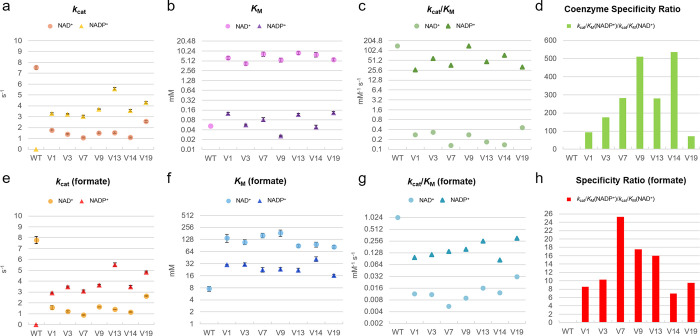
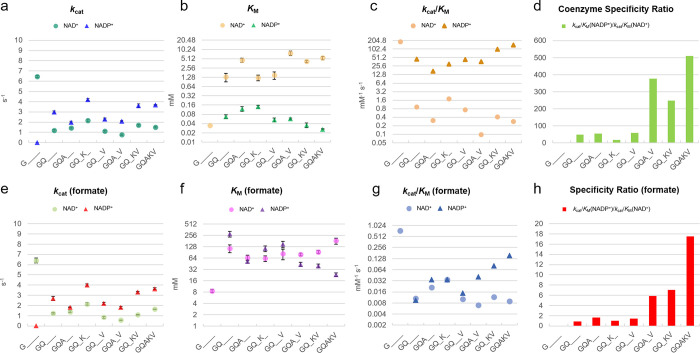
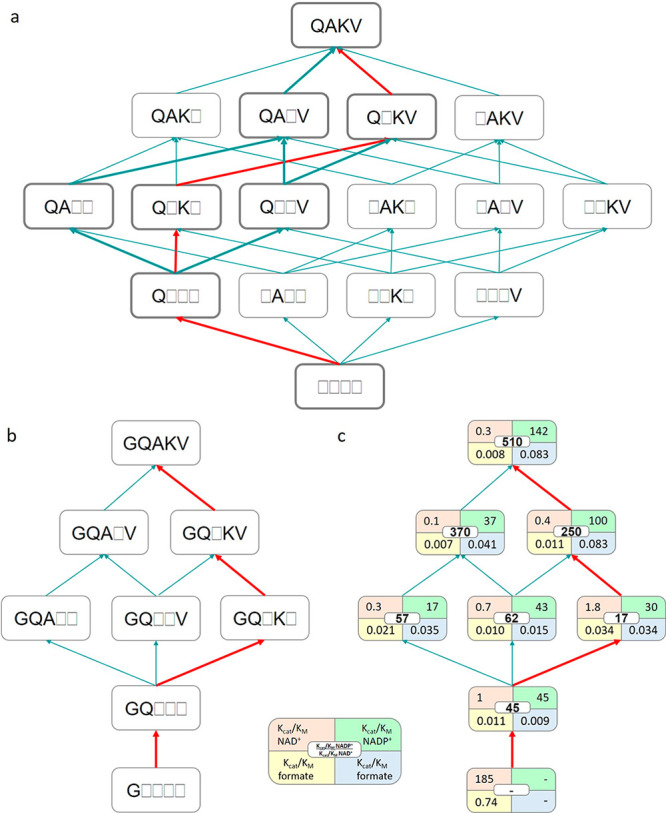
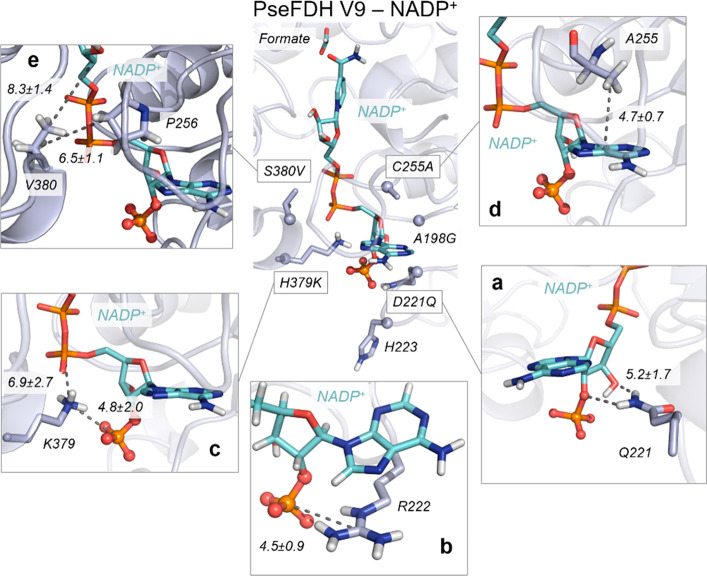
The efficient regeneration of cofactors is vital for the establishment of biocatalytic processes. Formate is an ideal electron donor for cofactor regeneration due to its general availability, low reduction potential, and benign byproduct (CO2). However, formate dehydrogenases (FDHs) are usually specific to NAD+, such that NADPH regeneration with formate is challenging. Previous studies reported naturally occurring FDHs or engineered FDHs that accept NADP+, but these enzymes show low kinetic efficiencies and specificities. Here, we harness the power of natural selection to engineer FDH variants to simultaneously optimize three properties: kinetic efficiency with NADP+, specificity toward NADP+, and affinity toward formate. By simultaneously mutating multiple residues of FDH from Pseudomonas sp. 101, which exhibits practically no activity toward NADP+, we generate a library of >106 variants. We introduce this library into an E. coli strain that cannot produce NADPH. By selecting for growth with formate as the sole NADPH source, we isolate several enzyme variants that support efficient NADPH regeneration. We find that the kinetically superior enzyme variant, harboring five mutations, has 5-fold higher efficiency and 14-fold higher specificity in comparison to the best enzyme previously engineered, while retaining high affinity toward formate. By using molecular dynamics simulations, we reveal the contribution of each mutation to the superior kinetics of this variant. We further determine how nonadditive epistatic effects improve multiple parameters simultaneously. Our work demonstrates the capacity of in vivo selection to identify highly proficient enzyme variants carrying multiple mutations which would be almost impossible to find using conventional screening methods.
Copyright © 2020 American Chemical Society.
Conflict of interest statement
The authors declare the following competing financial interest(s): A.B.-E. is a cofounder of b.fab, exploring the commercialization of microbial bioproduction using formate as a feedstock. The company was not involved in any way in performing or funding this study.
Figures

References
-
- Babel W. The Auxiliary Substrate Concept: From simple considerations to heuristically valuable knowledge. Eng. Life Sci. 2009, 9, 285–290. 10.1002/elsc.200900027. - DOI
LinkOut - more resources
Full Text Sources