

Flexible Usage and Interconnectivity of Diverse Cell Death Pathways Protect against Intracellular Infection
- PMID: 32735843
- PMCID: PMC7500851
- DOI: 10.1016/j.immuni.2020.07.004
Flexible Usage and Interconnectivity of Diverse Cell Death Pathways Protect against Intracellular Infection
Abstract
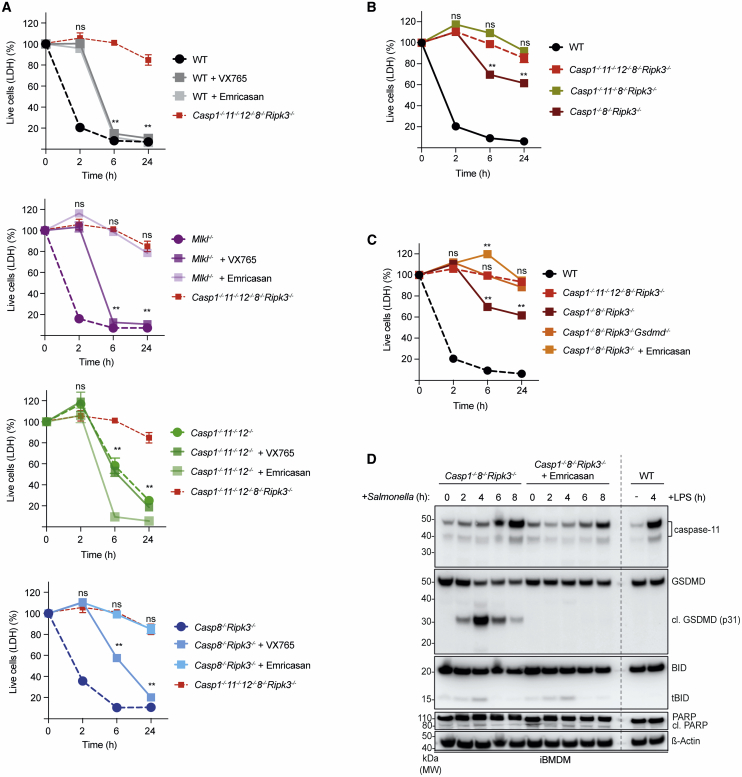
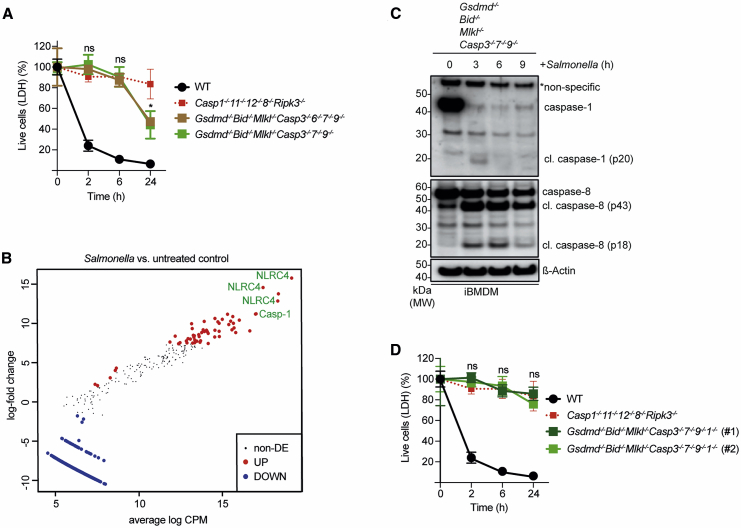
Programmed cell death contributes to host defense against pathogens. To investigate the relative importance of pyroptosis, necroptosis, and apoptosis during Salmonella infection, we infected mice and macrophages deficient for diverse combinations of caspases-1, -11, -12, and -8 and receptor interacting serine/threonine kinase 3 (RIPK3). Loss of pyroptosis, caspase-8-driven apoptosis, or necroptosis had minor impact on Salmonella control. However, combined deficiency of these cell death pathways caused loss of bacterial control in mice and their macrophages, demonstrating that host defense can employ varying components of several cell death pathways to limit intracellular infections. This flexible use of distinct cell death pathways involved extensive cross-talk between initiators and effectors of pyroptosis and apoptosis, where initiator caspases-1 and -8 also functioned as executioners when all known effectors of cell death were absent. These findings uncover a highly coordinated and flexible cell death system with in-built fail-safe processes that protect the host from intracellular infections.
Keywords: Salmonella; apoptosis; caspase-1; caspase-11; caspase-8; cell death; effector caspases; gasdermin D; necroptosis; pyroptosis.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures






Comment in
-
A Flexible and Deadly Way to Control Salmonella Infection.Immunity. 2020 Sep 15;53(3):471-473. doi: 10.1016/j.immuni.2020.08.009. Immunity. 2020. PMID: 32937145
References
-
- Alvarez-Diaz S., Dillon C.P., Lalaoui N., Tanzer M.C., Rodriguez D.A., Lin A., Lebois M., Hakem R., Josefsson E.C., O’Reilly L.A. The Pseudokinase MLKL and the Kinase RIPK3 Have Distinct Roles in Autoimmune Disease Caused by Loss of Death-Receptor-Induced Apoptosis. Immunity. 2016;45:513–526. - PMC - PubMed
-
- Aubrey B.J., Kelly G.L., Kueh A.J., Brennan M.S., O’Connor L., Milla L., Wilcox S., Tai L., Strasser A., Herold M.J. An inducible lentiviral guide RNA platform enables the identification of tumor-essential genes and tumor-promoting mutations in vivo. Cell Rep. 2015;10:1422–1432. - PubMed
-
- Bachem A., Makhlouf C., Binger K.J., de Souza D.P., Tull D., Hochheiser K., Whitney P.G., Fernandez-Ruiz D., Dahling S., Kastenmuller W. Microbiota-Derived Short-Chain Fatty Acids Promote the Memory Potential of Antigen-Activated CD8(+) T Cells. Immunity. 2019;51:285–297.e5. - PubMed
-
- Bedoui S., Kupz A., Wijburg O.L., Walduck A.K., Rescigno M., Strugnell R.A. Different bacterial pathogens, different strategies, yet the aim is the same: evasion of intestinal dendritic cell recognition. J. Immunol. 2010;184:2237–2242. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
