

Nanodisc scaffold peptide (NSPr) replaces detergent by reconstituting acyl-CoA:cholesterol acyltransferase 1 into peptidiscs
- PMID: 32735863
- PMCID: PMC7507747
- DOI: 10.1016/j.abb.2020.108518
Nanodisc scaffold peptide (NSPr) replaces detergent by reconstituting acyl-CoA:cholesterol acyltransferase 1 into peptidiscs
Abstract
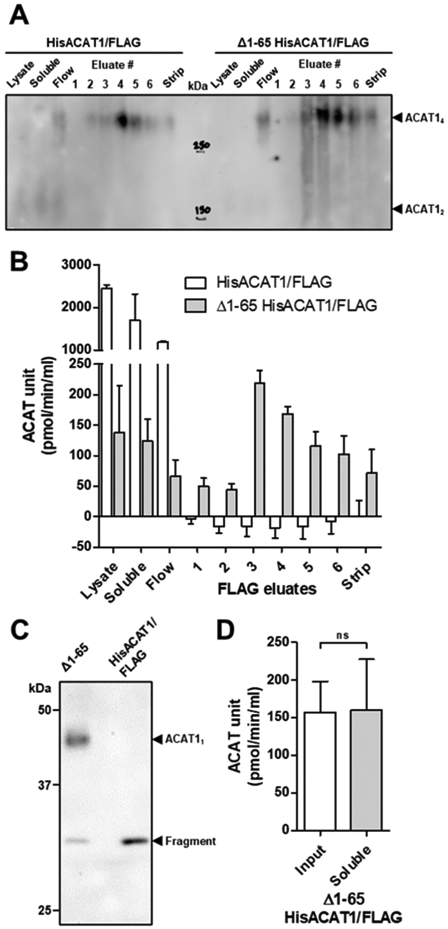
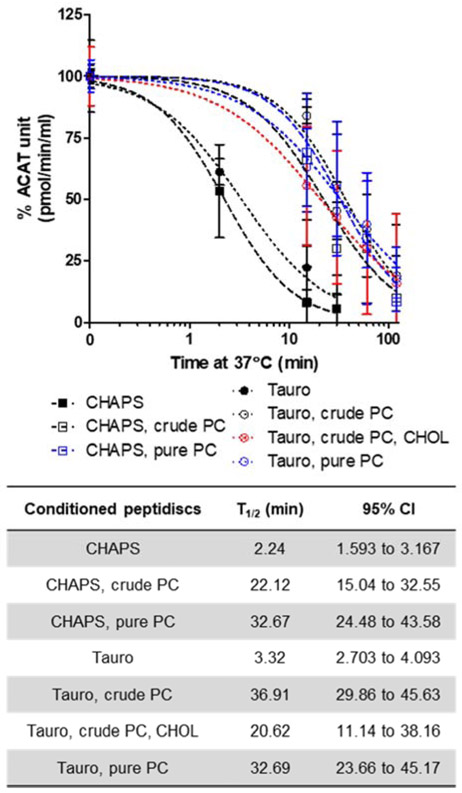
To conduct biochemical studies in vitro, membrane proteins (MPs) must be solubilized with detergents. While detergents are great tools, they can also inhibit the biological activity and/or perturb oligomerization of individual MPs. Nanodisc scaffold peptide (NSPr), an amphipathic peptide analog of ApoA1, was recently shown to reconstitute detergent solubilized MPs into peptidiscs in vitro. Acyl-coenzyme A:cholesterol acyltransferase 1 (ACAT1), also known as sterol O-acyltransferase 1 (SOAT1), plays a key role in cellular cholesterol storage in various cell types and is a drug target to treat multiple human diseases. ACAT1 contains nine transmembrane domains (TMDs) and primarily forms a homotetramer in vitro and in intact cells; deletion of the N-terminal dimerization domain produces a homodimer with full retention in catalytic activity. ACAT1 is prone to inactivation by numerous detergents. Here we pursued the use of NSPr to overcome the detergent-induced inactivation of ACAT1 by generating near detergent-free ACAT1 peptidiscs. Based on native-PAGE analysis, we showed that NSPr reconstitutes ACAT1 into soluble peptidiscs, in which ACAT1 exists predominantly in oligomeric states greater than a homotetramer. The formation of these higher-order oligomeric states was independent of the N-terminal dimerization domain, suggesting that the oligomerization is mediated through hydrophobic interactions of multiple ACAT1 subunits. ACAT1 peptidiscs were still susceptible to heat-mediated inactivation, presumably due to the residual detergent (CHAPS) bound to ACAT1. We then conditioned ACAT1 with phosphatidylcholine (PC) to replace CHAPS prior to the formation of ACAT1 peptidiscs. The results showed, when PC was included, ACAT1 was present mainly in higher-order oligomeric states with greater enzymatic activity. With PC present, the enzymatic activity of ACAT1 peptidiscs was protected from heat-mediated inactivation. These results support the use of NSPr to create a near detergent-free solution of ACAT1 in peptidiscs for various in vitro studies. Our current results also raise the possibility that, under certain conditions, ACAT1 may form higher-order oligomeric states in vivo.
Keywords: ACAT/SOAT; Detergent-free; Membrane protein; Oligomerization; Peptidisc; Phospholipid.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest
The authors declare that they have no conflicts of interest with the contents of this article.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
