

Quality Control of ER Membrane Proteins by the RNF185/Membralin Ubiquitin Ligase Complex
- PMID: 32738194
- PMCID: PMC7482433
- DOI: 10.1016/j.molcel.2020.07.009
Quality Control of ER Membrane Proteins by the RNF185/Membralin Ubiquitin Ligase Complex
Erratum in
-
Quality Control of ER Membrane Proteins by the RNF185/Membralin Ubiquitin Ligase Complex.Mol Cell. 2020 Oct 15;80(2):374-375. doi: 10.1016/j.molcel.2020.09.023. Mol Cell. 2020. PMID: 33065021 Free PMC article. No abstract available.
Abstract
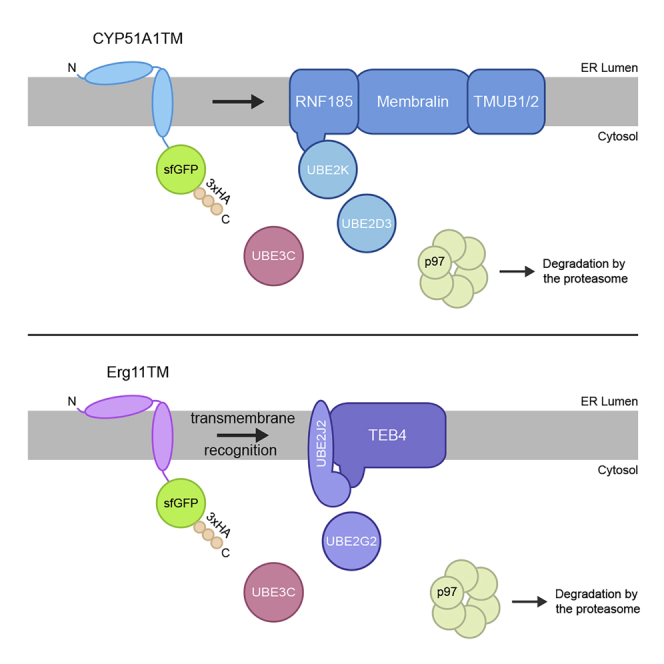
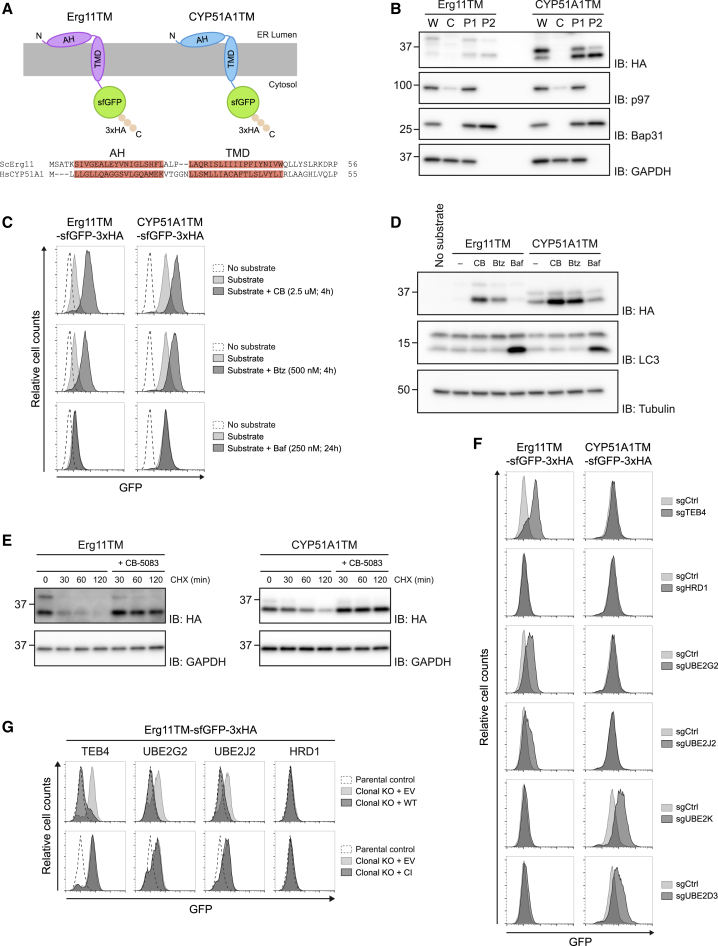
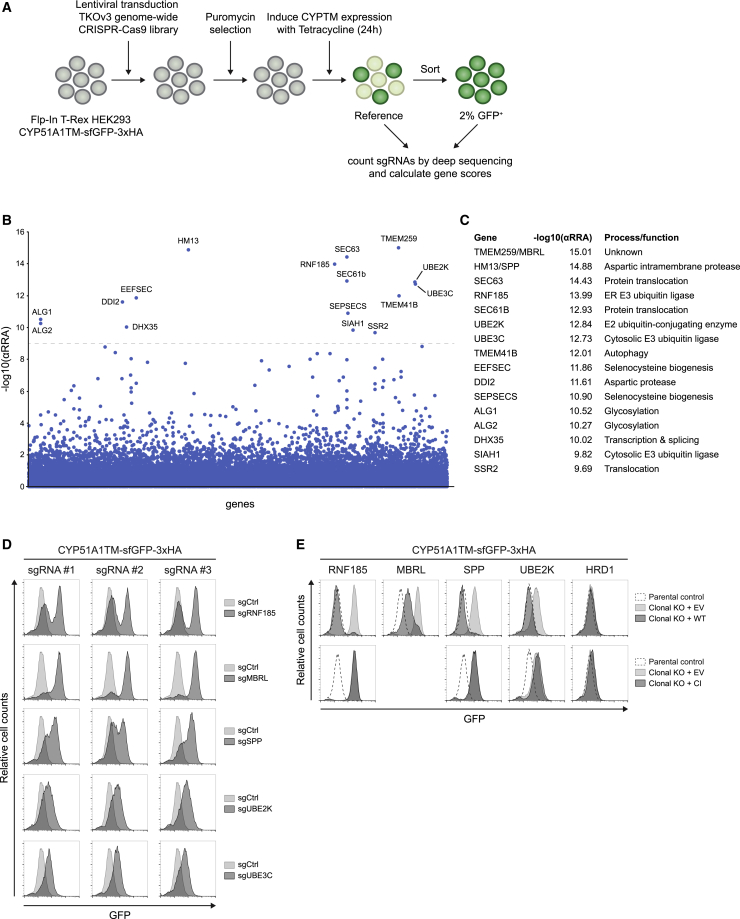
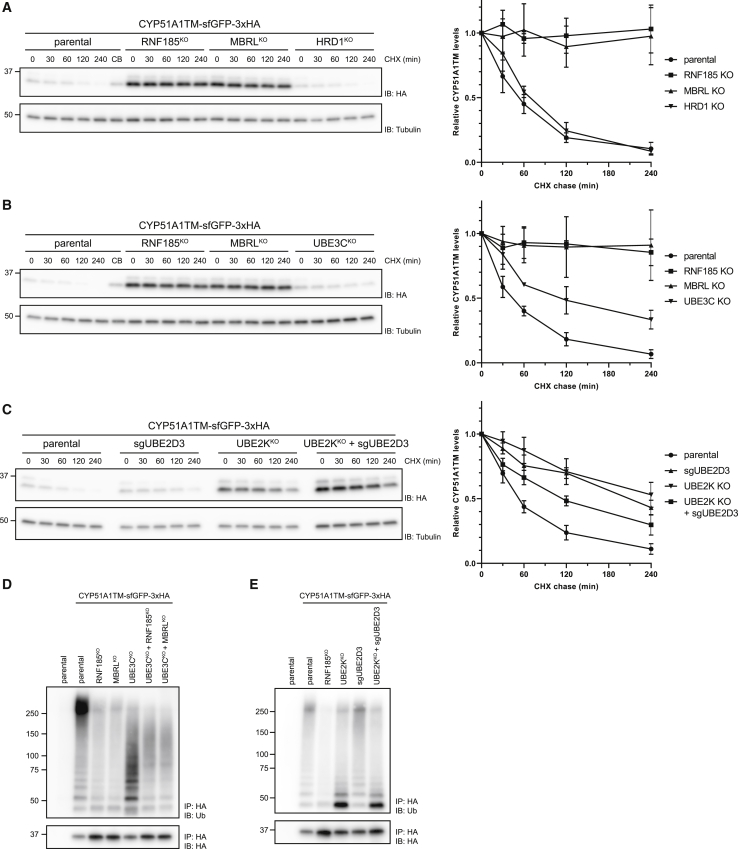
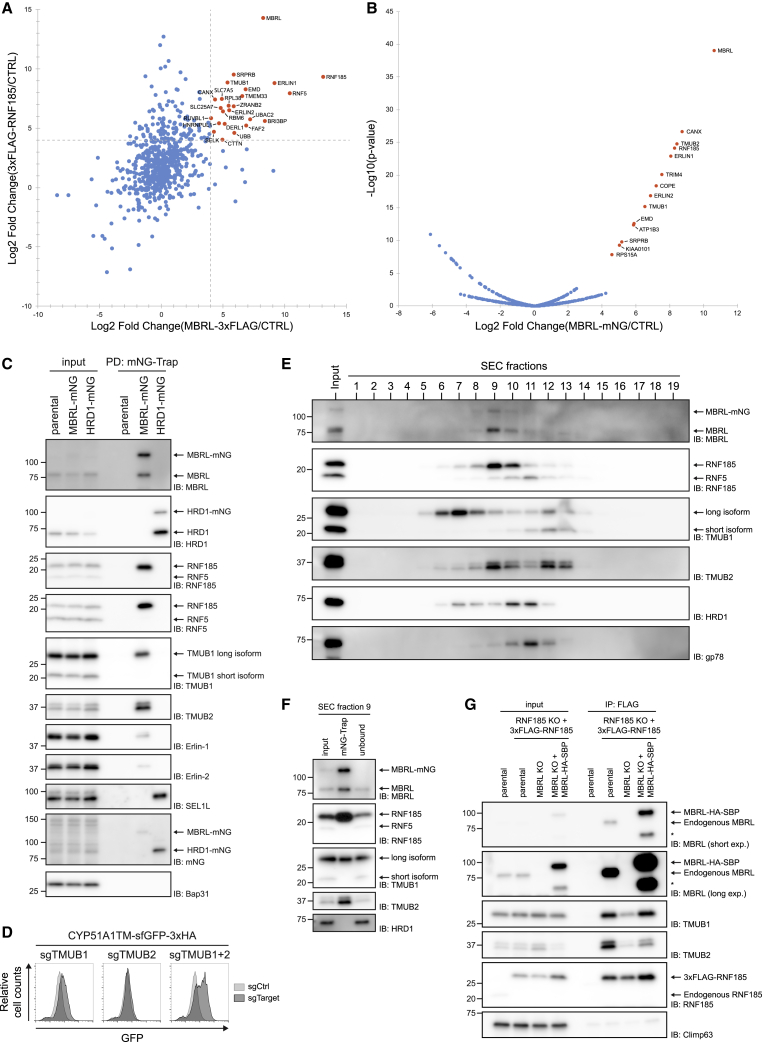
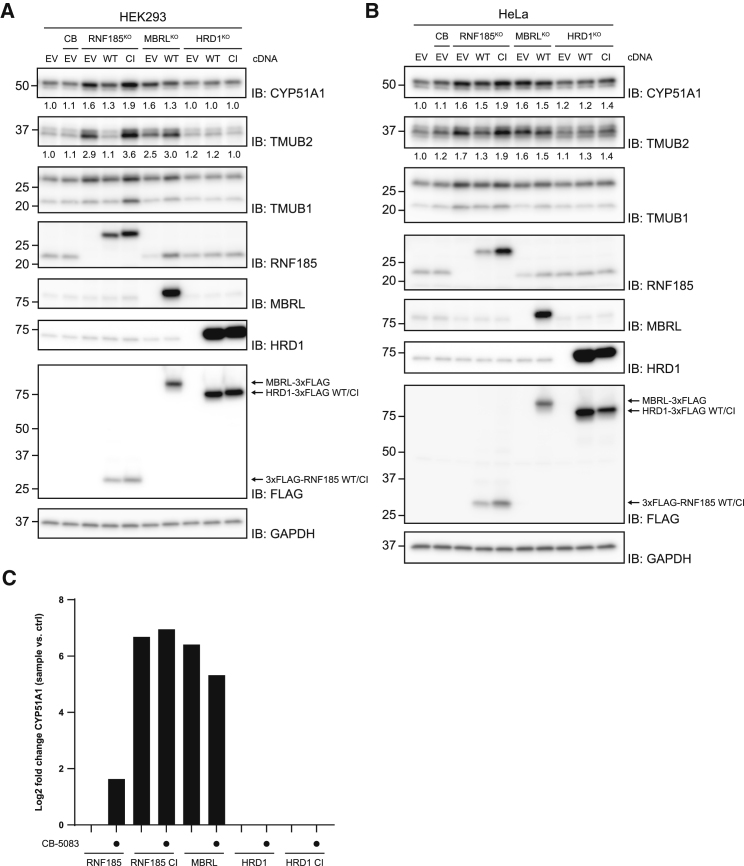
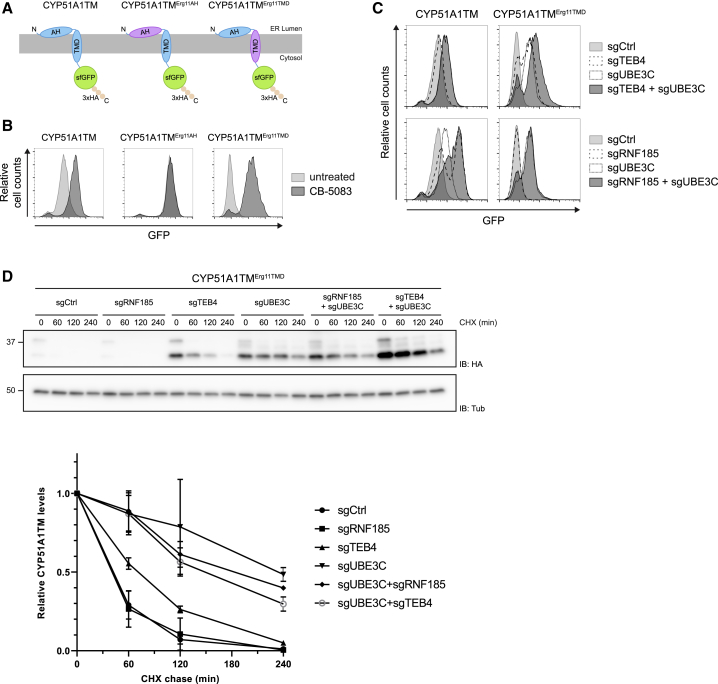
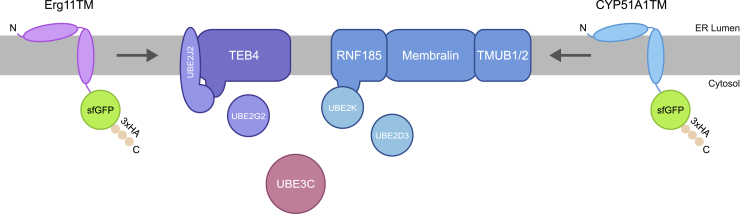
Misfolded proteins in the endoplasmic reticulum (ER) are degraded by ER-associated degradation (ERAD). Although ERAD components involved in degradation of luminal substrates are well characterized, much less is known about quality control of membrane proteins. Here, we analyzed the degradation pathways of two short-lived ER membrane model proteins in mammalian cells. Using a CRISPR-Cas9 genome-wide library screen, we identified an ERAD branch required for quality control of a subset of membrane proteins. Using biochemical and mass spectrometry approaches, we showed that this ERAD branch is defined by an ER membrane complex consisting of the ubiquitin ligase RNF185, the ubiquitin-like domain containing proteins TMUB1/2 and TMEM259/Membralin, a poorly characterized protein. This complex cooperates with cytosolic ubiquitin ligase UBE3C and p97 ATPase in degrading their membrane substrates. Our data reveal that ERAD branches have remarkable specificity for their membrane substrates, suggesting that multiple, perhaps combinatorial, determinants are involved in substrate selection.
Keywords: ER-associated degradation; ERAD; RNF185; TEB4/MARCH6; TMEM259; TMUB1/TMUB2; UBE3C; endoplasmic reticulum; membralin; protein quality control.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Andersson O., von Euler G. Characterization and expression of the gene encoding membralin, an evolutionary conserved protein expressed in the central nervous system. Brain Res. Gene Expr. Patterns. 2002;1:205–212. - PubMed
-
- Carvalho P., Goder V., Rapoport T.A. Distinct ubiquitin-ligase complexes define convergent pathways for the degradation of ER proteins. Cell. 2006;126:361–373. - PubMed
-
- Chen Z., Pickart C.M. A 25-kilodalton ubiquitin carrier protein (E2) catalyzes multi-ubiquitin chain synthesis via lysine 48 of ubiquitin. J. Biol. Chem. 1990;265:21835–21842. - PubMed
-
- Chen Y.-C., Davidson B., Cheng C.-C., Maitra A., Giuntoli R.L., 2nd, Hruban R.H., Wang T.-L., Shih IeM. Identification and characterization of membralin, a novel tumor-associated gene, in ovarian carcinoma. Biochim. Biophys. Acta. 2005;1730:96–102. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
