

Single-cell transcriptomics of human islet ontogeny defines the molecular basis of β-cell dedifferentiation in T2D
- PMID: 32739450
- PMCID: PMC7471622
- DOI: 10.1016/j.molmet.2020.101057
Single-cell transcriptomics of human islet ontogeny defines the molecular basis of β-cell dedifferentiation in T2D
Abstract
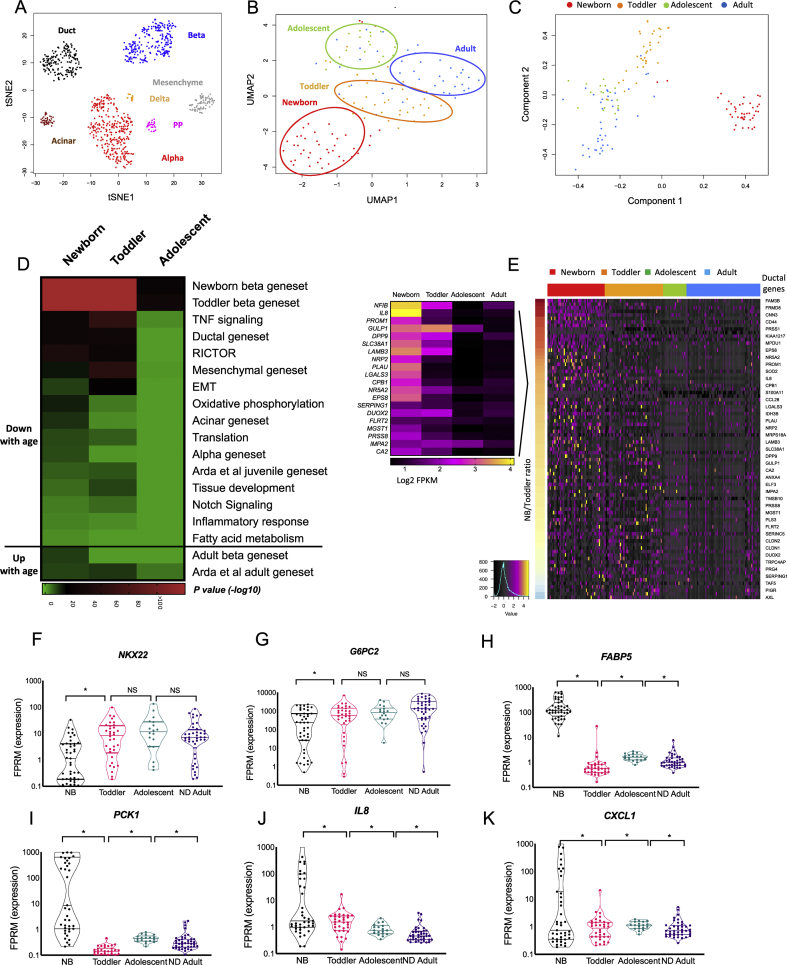
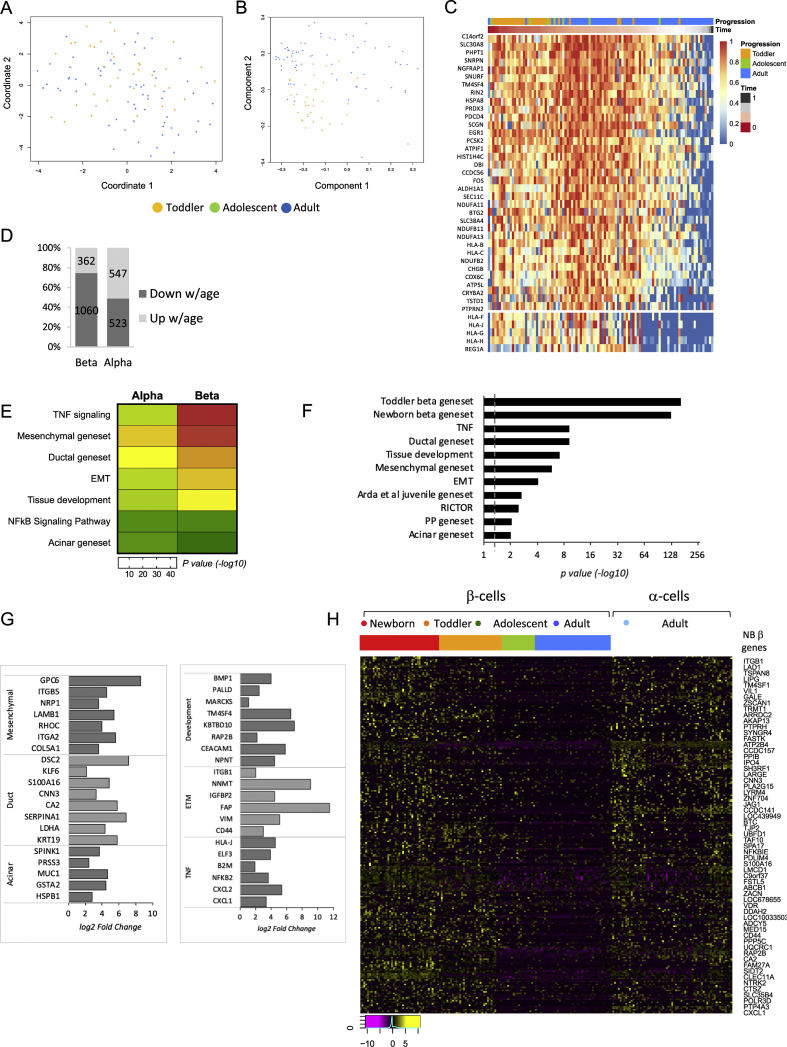
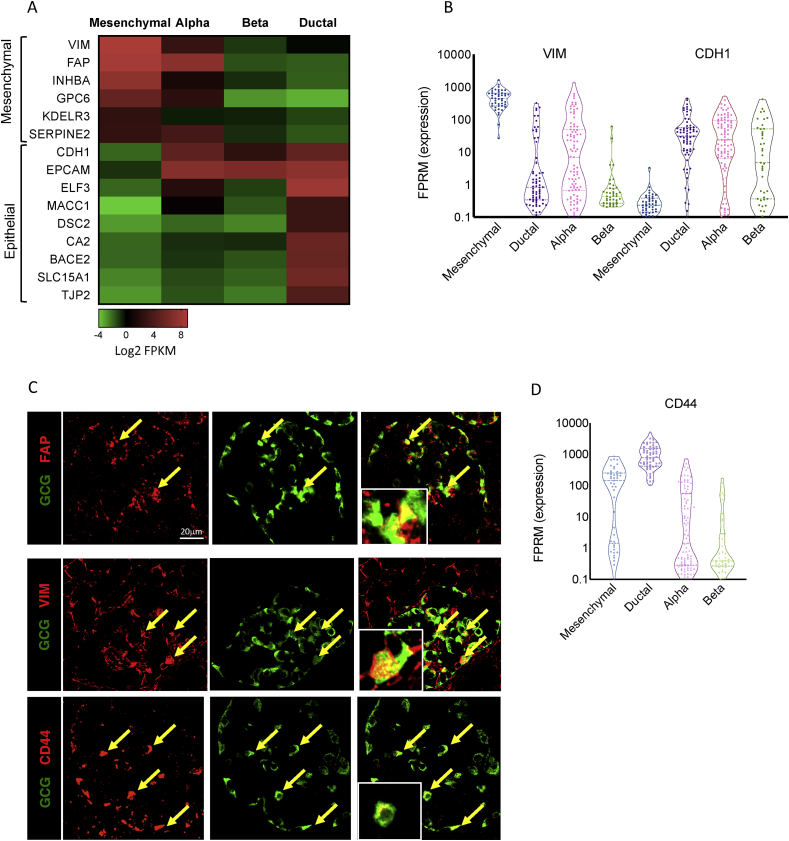
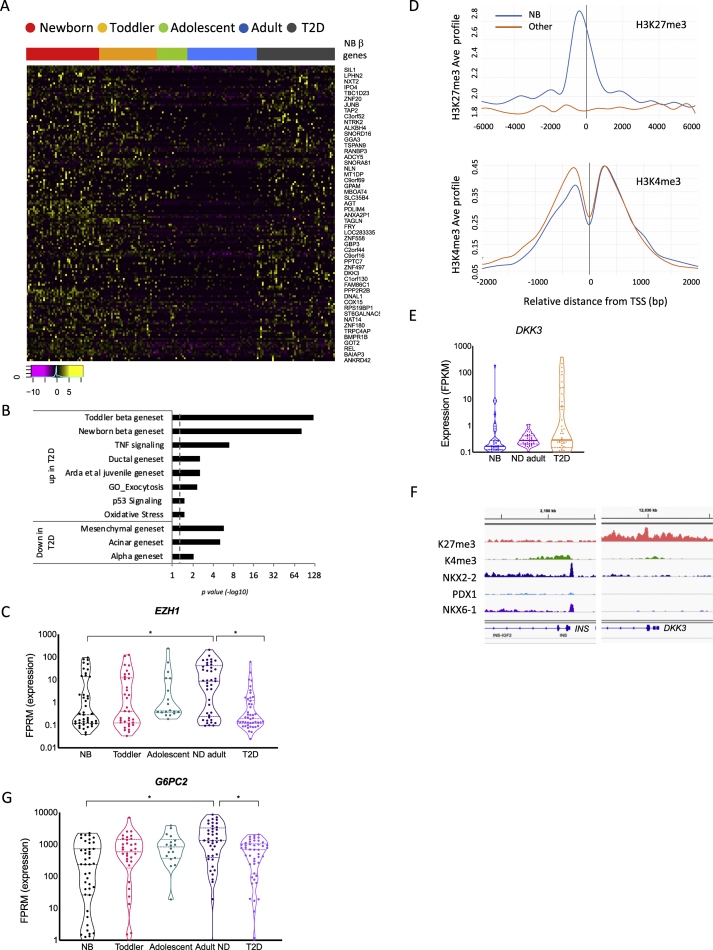
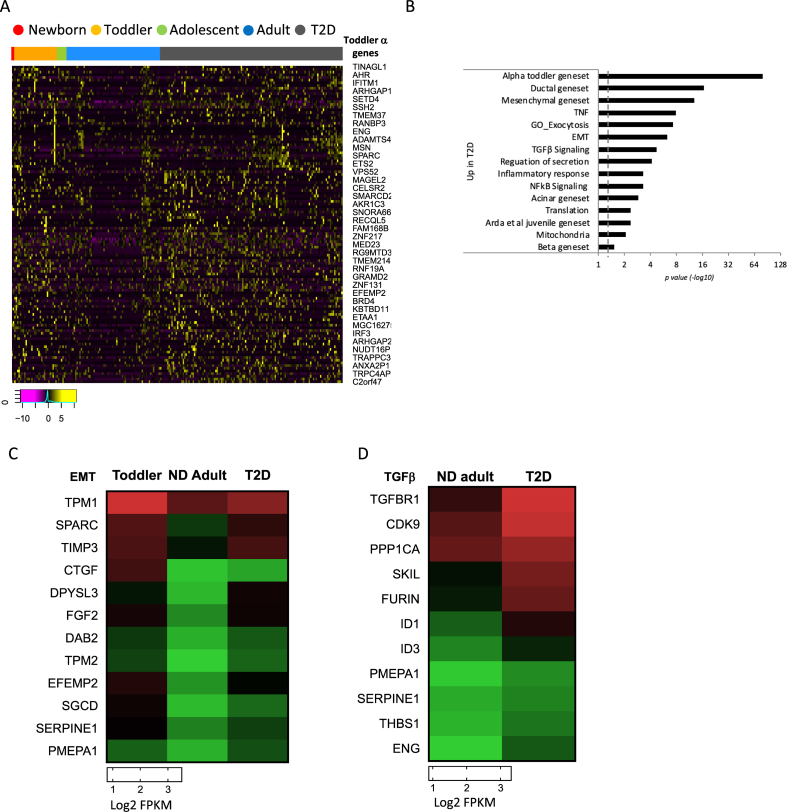
Objective: Dedifferentiation of pancreatic β-cells may reduce islet function in type 2 diabetes (T2D). However, the prevalence, plasticity and functional consequences of this cellular state remain unknown.
Methods: We employed single-cell RNAseq to detail the maturation program of α- and β-cells during human ontogeny. We also compared islets from non-diabetic and T2D individuals.
Results: Both α- and β-cells mature in part by repressing non-endocrine genes; however, α-cells retain hallmarks of an immature state, while β-cells attain a full β-cell specific gene expression program. In islets from T2D donors, both α- and β-cells have a less mature expression profile, de-repressing the juvenile genetic program and exocrine genes and increasing expression of exocytosis, inflammation and stress response signalling pathways. These changes are consistent with the increased proportion of β-cells displaying suboptimal function observed in T2D islets.
Conclusions: These findings provide new insights into the molecular program underlying islet cell maturation during human ontogeny and the loss of transcriptomic maturity that occurs in islets of type 2 diabetics.
Keywords: Human islet; Ontogeny; Single cell RNAseq; Type 2 diabetes; α-Cell; β-Cell.
Copyright © 2020 The Author(s). Published by Elsevier GmbH.. All rights reserved.
Figures
References
-
- Donath M.Y., Ehses J.A., Maedler K., Schumann D.M., Ellingsgaard H., Eppler E. Mechanisms of beta-cell death in type 2 diabetes. Diabetes. 2005;54(Suppl 2):S108–S113. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
