

HFE inhibits type I IFNs signaling by targeting the SQSTM1-mediated MAVS autophagic degradation
- PMID: 32746697
- PMCID: PMC8386699
- DOI: 10.1080/15548627.2020.1804683
HFE inhibits type I IFNs signaling by targeting the SQSTM1-mediated MAVS autophagic degradation
Erratum in
-
Correction.Autophagy. 2025 Jun;21(6):ii-iii. doi: 10.1080/15548627.2022.2122346. Epub 2022 Oct 12. Autophagy. 2025. PMID: 36224706 Free PMC article. No abstract available.
Abstract
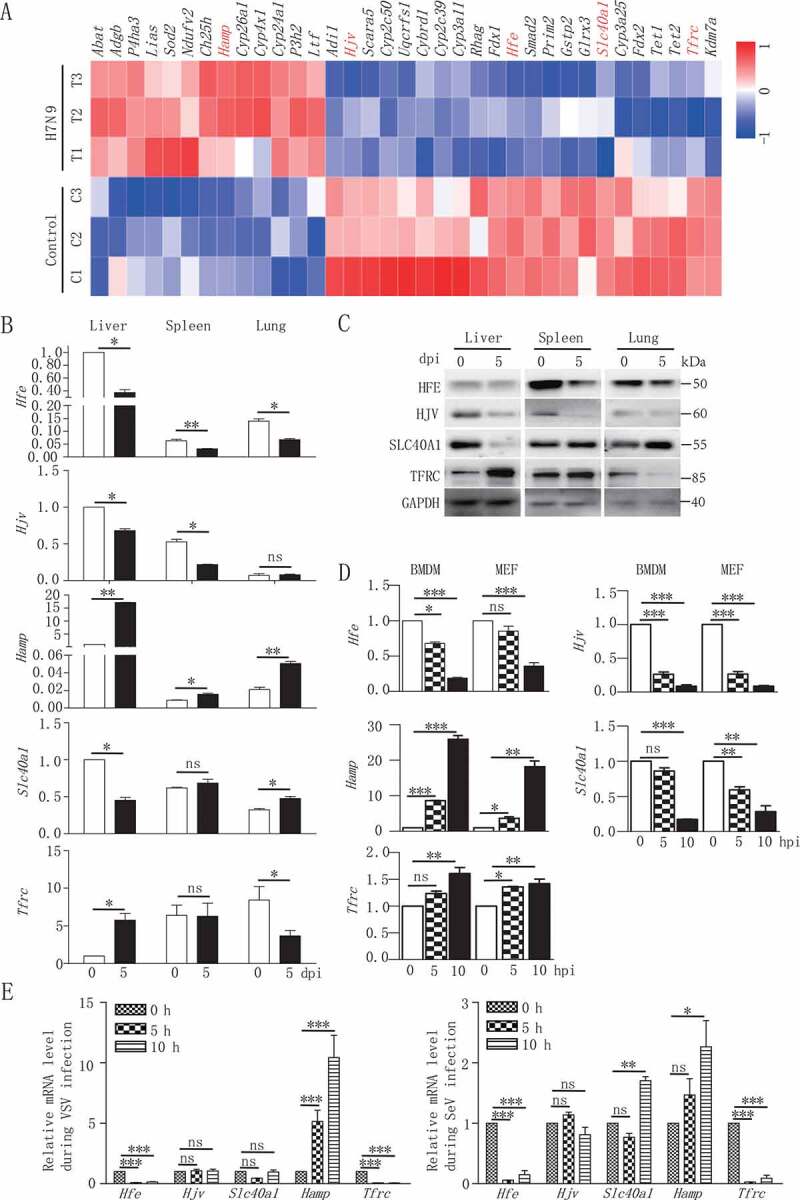
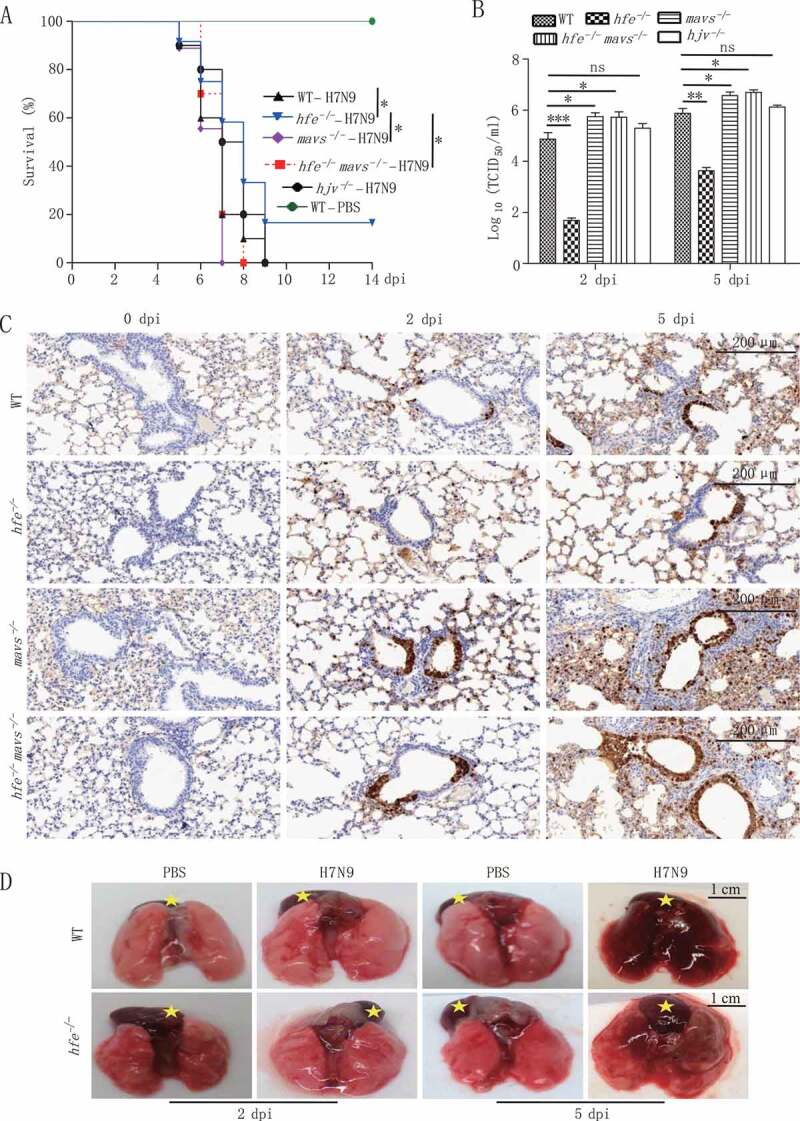
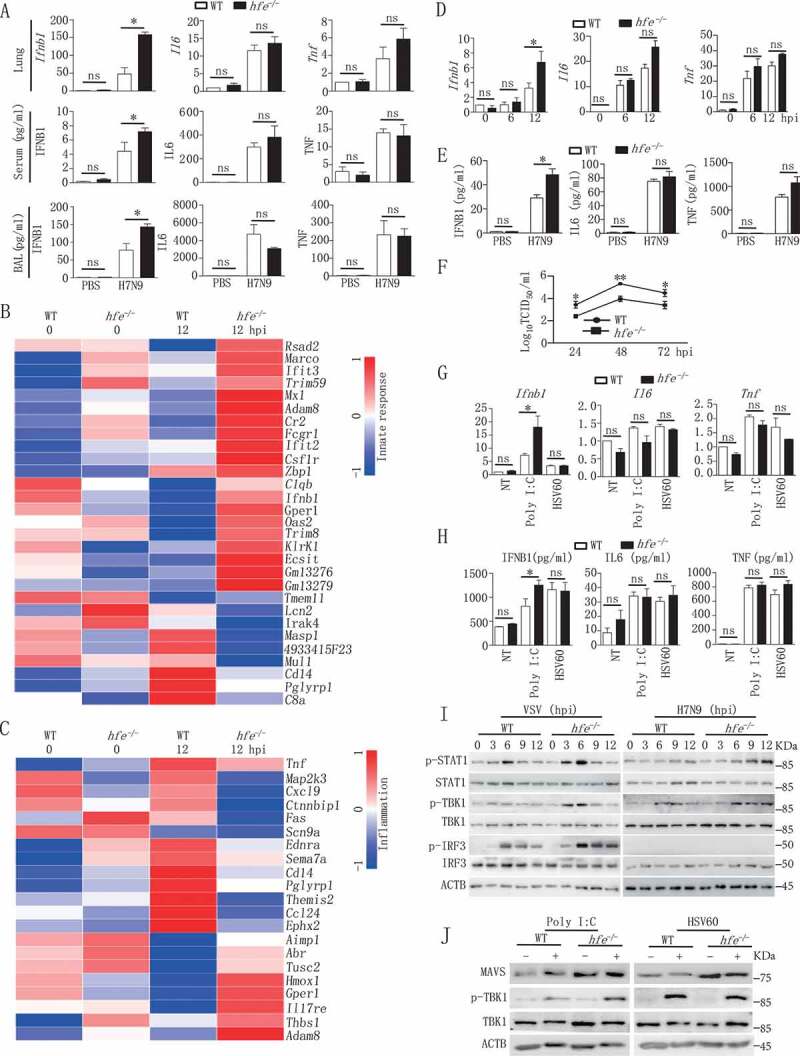
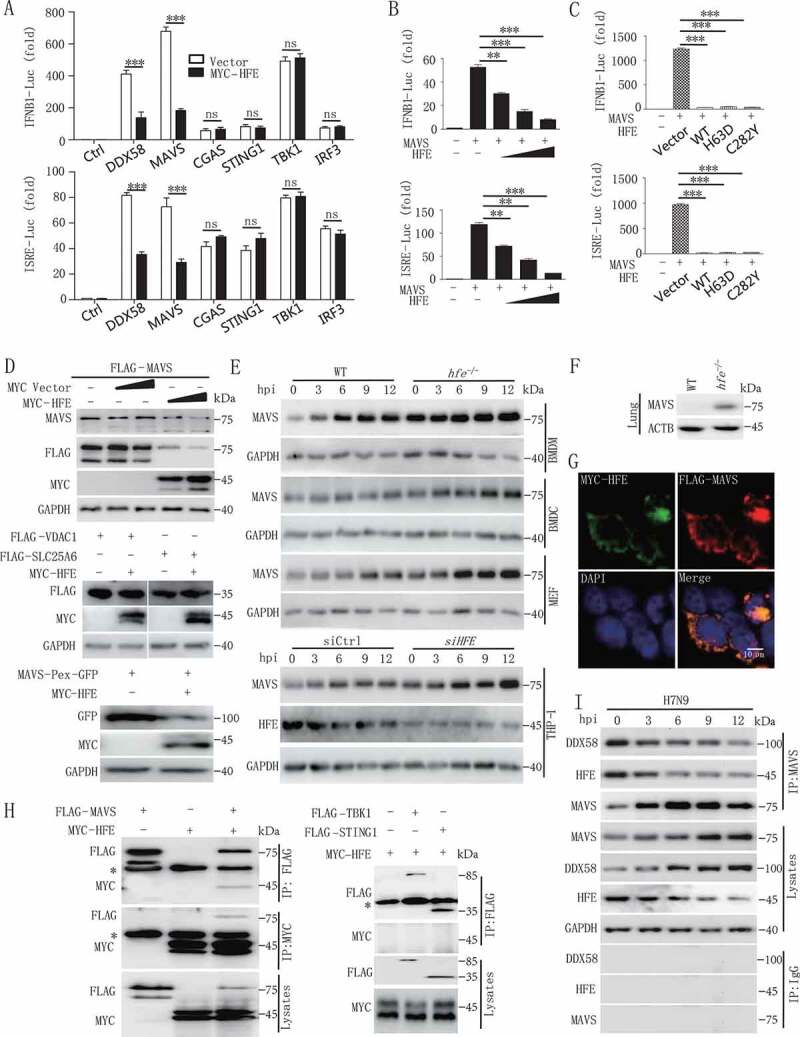
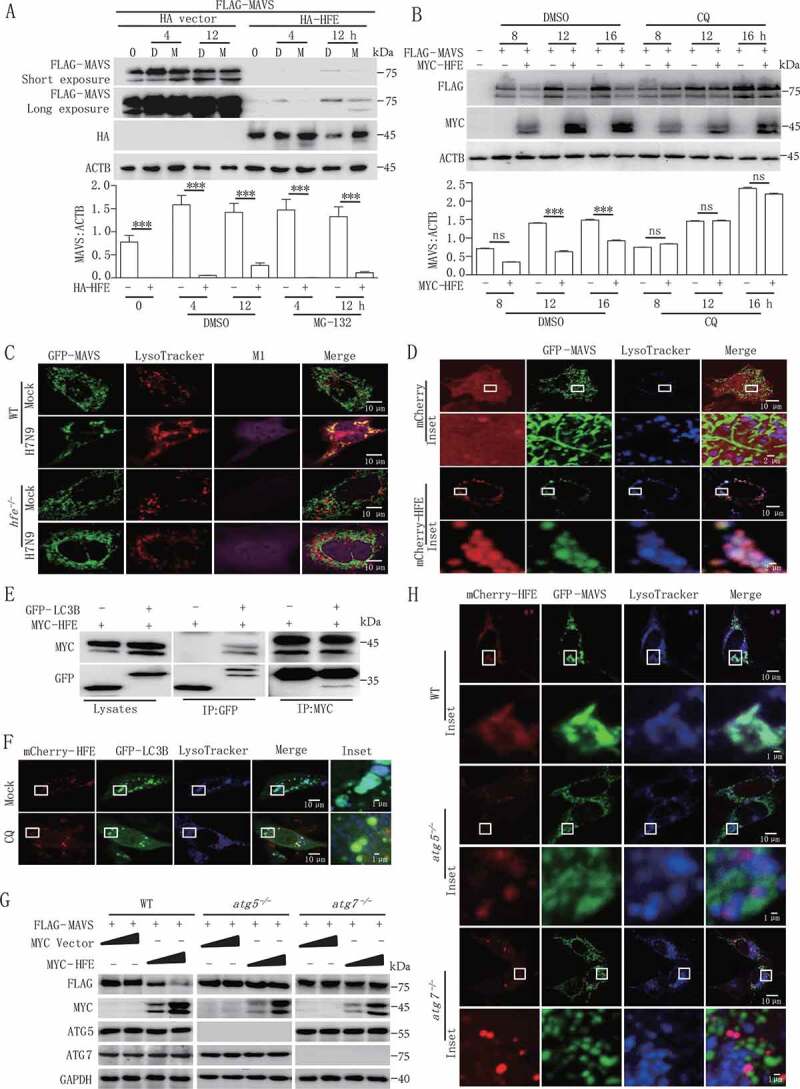
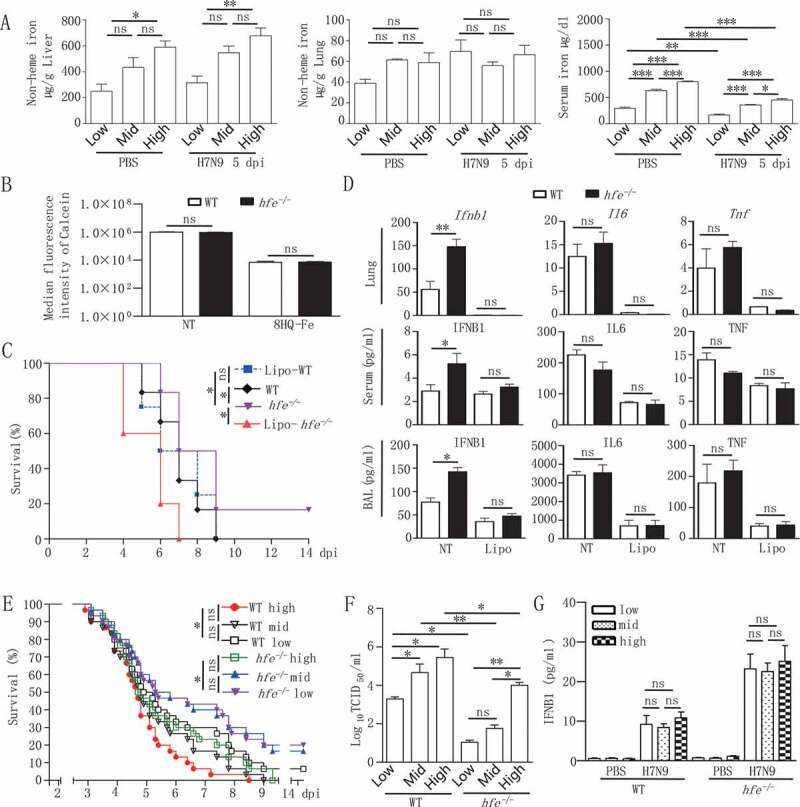
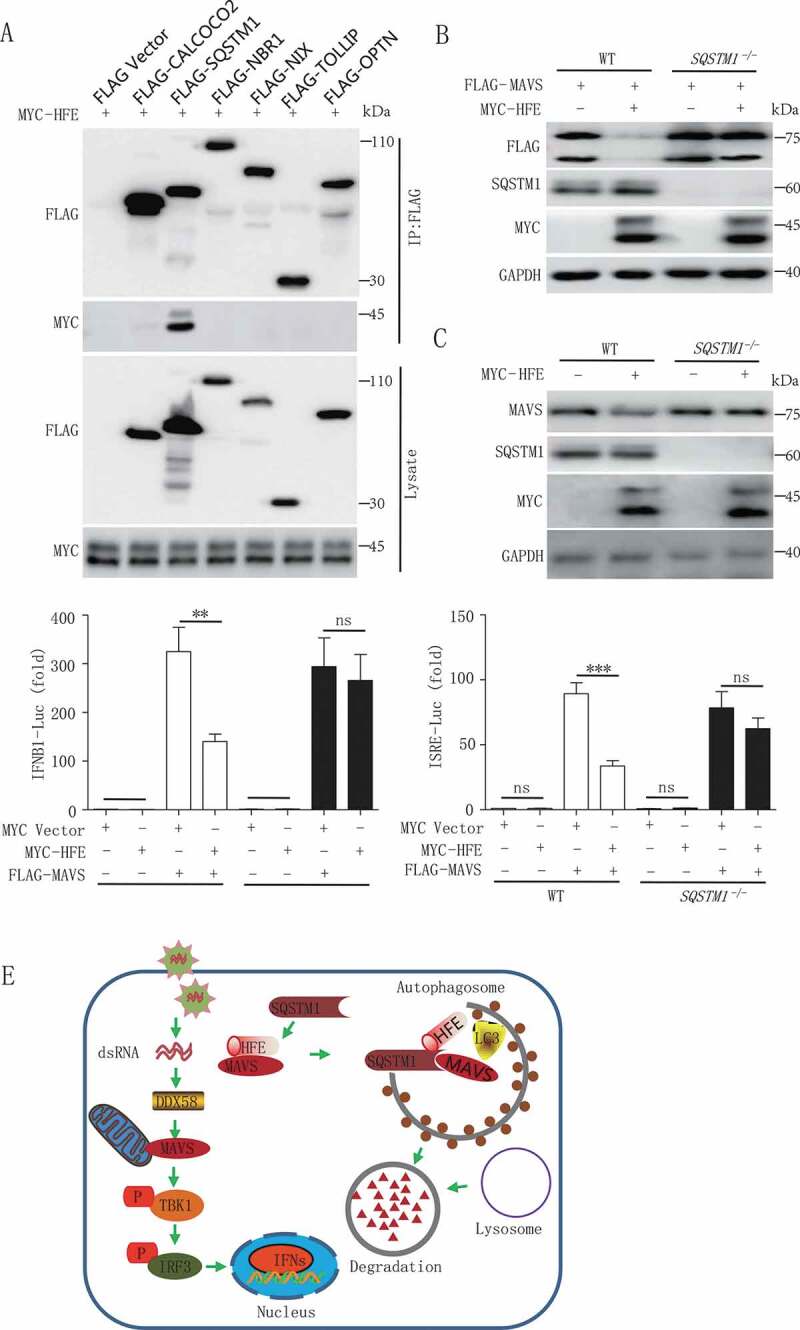
Iron metabolism is involved in numerous physiological processes such as erythropoiesis, oxidative metabolism. However, the in vivo physiological functions of the iron metabolism-related gene Hfe in immune response during viral infection remain poorly understood. Here, we identified 5 iron metabolism-associated genes specifically affected during RNA virus infection by a high-throughput assay and further found that HFE was a key negative regulator of RIG-I-like receptors (RLR)-mediated type I interferons (IFNs) signaling. RNA virus infection inhibited the binding of HFE to MAVS (mitochondrial antiviral signaling protein) and blocked MAVS degradation via selective autophagy. HFE mediated MAVS autophagic degradation by binding to SQSTM1/p62. Depletion of Hfe abrogated the autophagic degradation of MAVS, leading to the stronger antiviral immune response. These findings established a novel regulatory role of selective autophagy in innate antiviral immune response by the iron metabolism-related gene Hfe. These data further provided insights into the crosstalk among iron metabolism, autophagy, and innate immune response.Abbreviations: ATG: autophagy-related; BAL: bronchoalveolar lavage fluid; BMDMs: bone marrow-derived macrophages; CGAS: cyclic GMP-AMP synthase; CQ: chloroquine; Dpi: days post-infection; ELISA: enzyme-linked immunosorbent assay; GFP: green fluorescent protein; HAMP: hepcidin antimicrobial peptide; Hpi: hours post-infection; HJV: hemojuvelin BMP co-receptor; IFNs: interferons; IL6: interleukin 6; IRF3: interferon regulatory factor 3; ISRE: interferon-stimulated response element; Lipo: clodronate liposomes; MAP1LC3B/LC3B: microtubule-associated protein 1 light chain 3 beta; MAVS: mitochondrial antiviral signaling protein; MEFs: mouse embryonic fibroblasts; SLC40A1/FPN1: solute carrier family 40 (iron-regulated transporter), member 1; flatiron; SQSTM1/p62: sequestosome 1; STAT1: signal transducer and activator of transcription 1; STING1/STING: stimulator of interferon response cGAMP interactor 1; TBK1: TANK-binding kinase 1; TFRC/TfR1: transferrin receptor; TNF/TNFα: tumor necrosis factor; WT: wild type.
Keywords: Autophagy; HFE; MAVS degradation; RNA virus; innate immune; iron metabolism.
Conflict of interest statement
No potential conflict of interest was reported by the authors.
Figures
References
-
- Ramalingam TS, West AP Jr., Lebron JA, et al. Binding to the transferrin receptor is required for endocytosis of HFE and regulation of iron homeostasis. Nat Cell Biol. 2000;2(12):953–957. - PubMed
-
- Roy CN, Carlson EJ, Anderson EL, et al. Interactions of the ectodomain of HFE with the transferrin receptor are critical for iron homeostasis in cells. FEBS Lett. 2000;484(3):271–274.. - PubMed
-
- Salter-Cid L, Peterson PA, Yang Y. The major histocompatibility complex-encoded HFE in iron homeostasis and immune function. Immunol Res. 2000;22(1):43–59. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous