

Human pluripotent stem cell-derived cardiomyocytes as a target platform for paracrine protection by cardiac mesenchymal stromal cells
- PMID: 32747668
- PMCID: PMC7400574
- DOI: 10.1038/s41598-020-69495-w
Human pluripotent stem cell-derived cardiomyocytes as a target platform for paracrine protection by cardiac mesenchymal stromal cells
Abstract
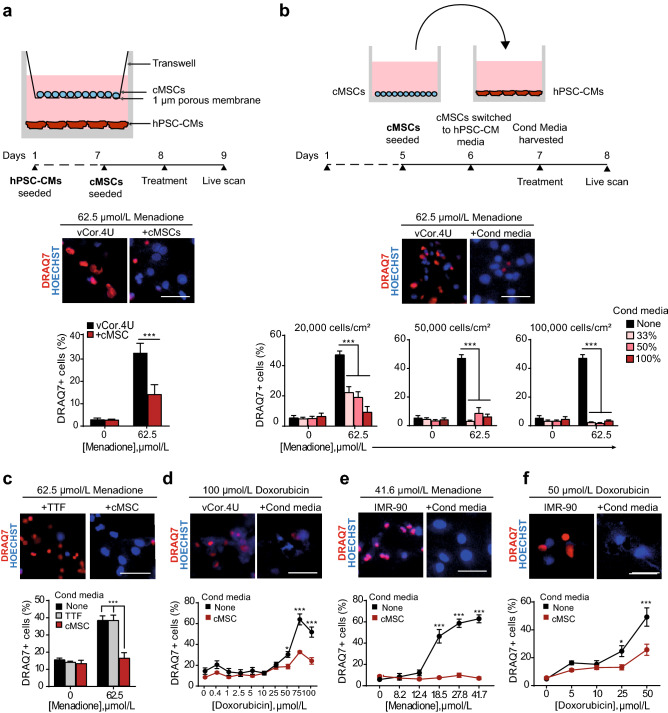
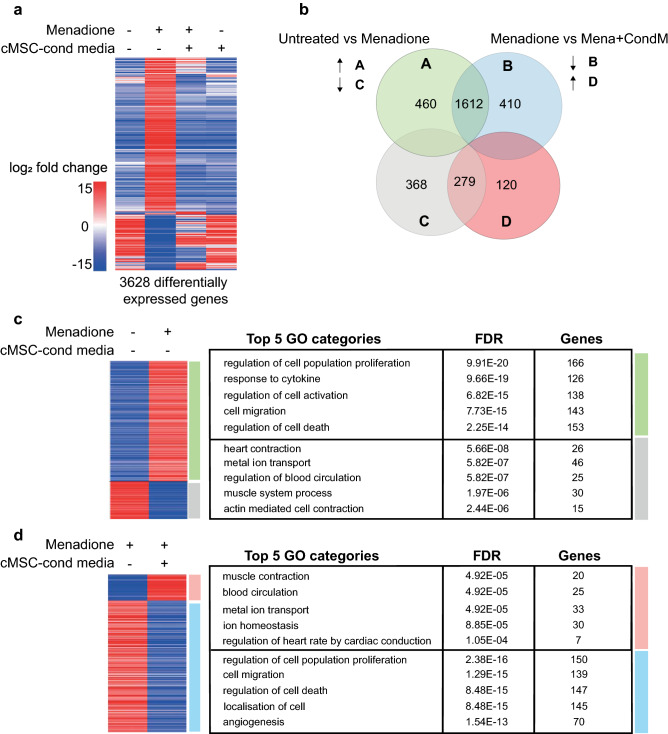
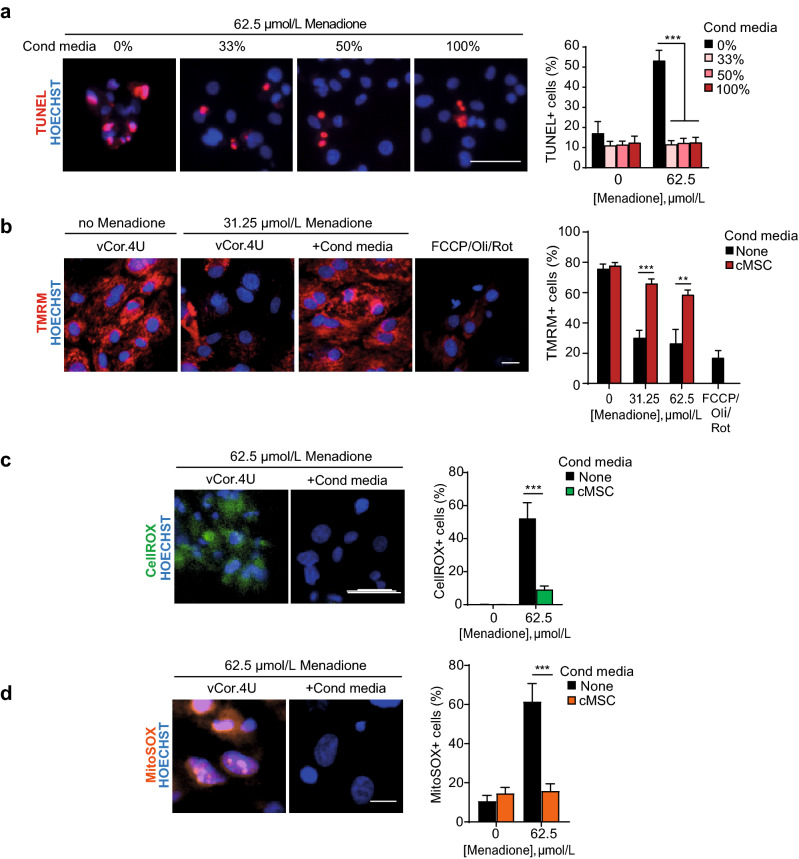
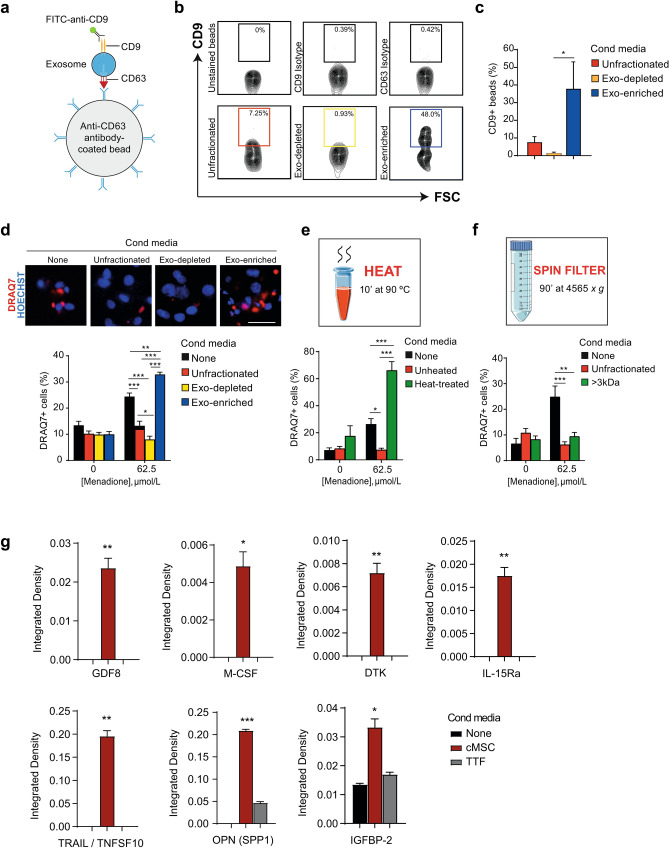
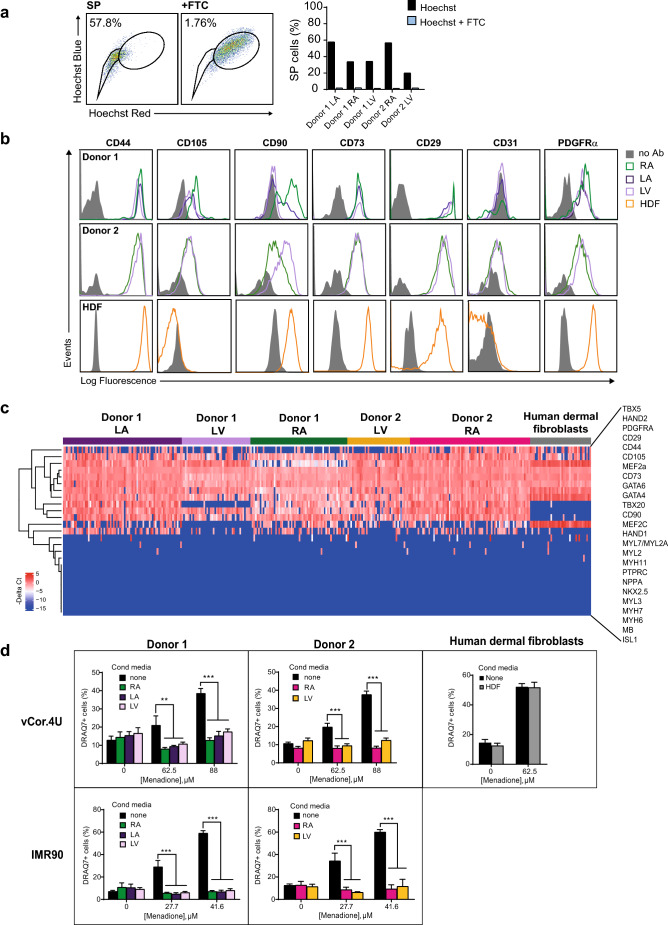
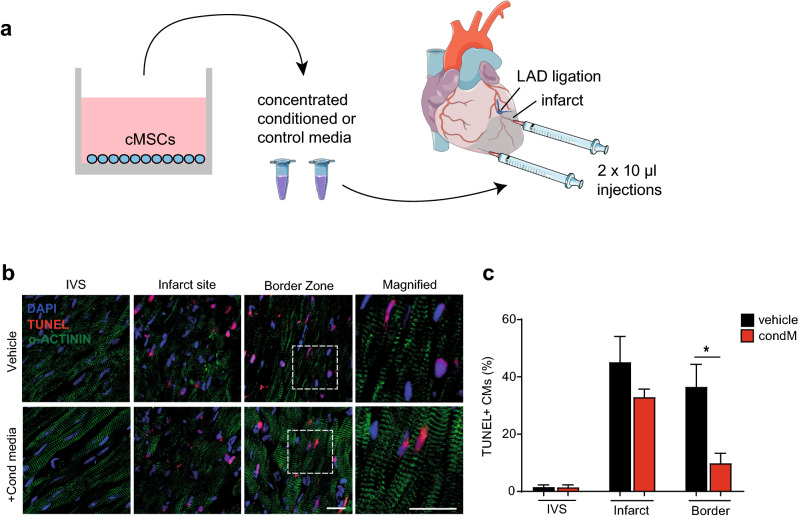
Ischemic heart disease remains the foremost cause of death globally, with survivors at risk for subsequent heart failure. Paradoxically, cell therapies to offset cardiomyocyte loss after ischemic injury improve long-term cardiac function despite a lack of durable engraftment. An evolving consensus, inferred preponderantly from non-human models, is that transplanted cells benefit the heart via early paracrine signals. Here, we tested the impact of paracrine signals on human cardiomyocytes, using human pluripotent stem cell-derived cardiomyocytes (hPSC-CMs) as the target of mouse and human cardiac mesenchymal stromal cells (cMSC) with progenitor-like features. In co-culture and conditioned medium studies, cMSCs markedly inhibited human cardiomyocyte death. Little or no protection was conferred by mouse tail tip or human skin fibroblasts. Consistent with the results of transcriptomic profiling, functional analyses showed that the cMSC secretome suppressed apoptosis and preserved cardiac mitochondrial transmembrane potential. Protection was independent of exosomes under the conditions tested. In mice, injecting cMSC-conditioned media into the infarct border zone reduced apoptotic cardiomyocytes > 70% locally. Thus, hPSC-CMs provide an auspicious, relevant human platform to investigate extracellular signals for cardiac muscle survival, substantiating human cardioprotection by cMSCs, and suggesting the cMSC secretome or its components as potential cell-free therapeutic products.
Conflict of interest statement
R.P.S. and A.M.C. were employees of Kardia Therapeutics. R.P.S. and M.D.S. were founders and stock-holders. All other authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
