

Development of Noonan syndrome by deregulation of allosteric SOS autoactivation
- PMID: 32753483
- PMCID: PMC7521655
- DOI: 10.1074/jbc.RA120.013275
Development of Noonan syndrome by deregulation of allosteric SOS autoactivation
Abstract
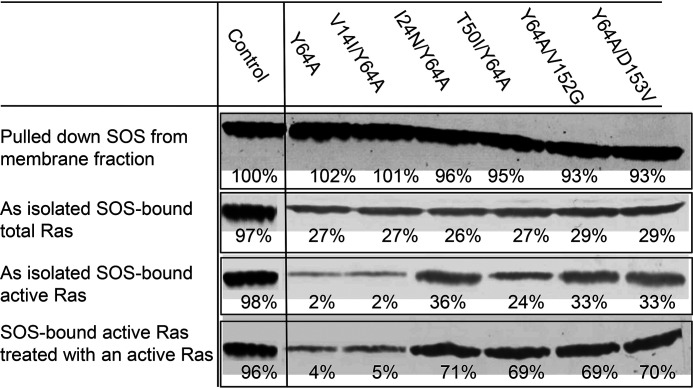
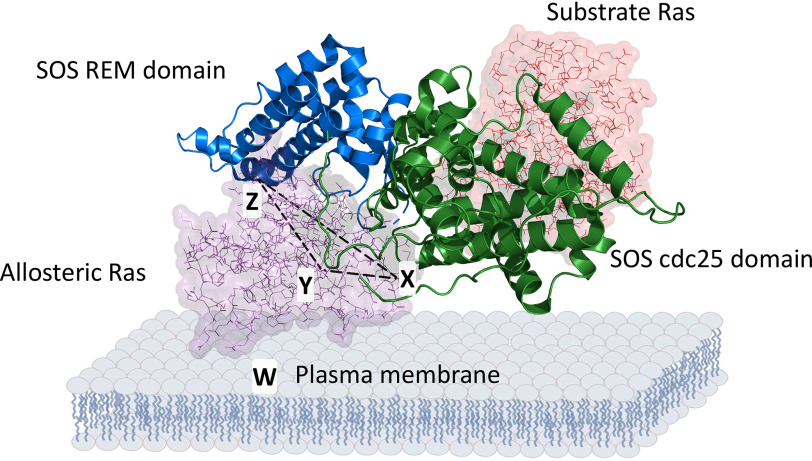
Ras family proteins play an essential role in several cellular functions, including growth, differentiation, and survival. The mechanism of action of Ras mutants in Costello syndrome and cancers has been identified, but the contribution of Ras mutants to Noonan syndrome, a genetic disorder that prevents normal development in various parts of the body, is unknown. Son of Sevenless (SOS) is a Ras guanine nucleotide exchange factor. In response to Ras-activating cell signaling, SOS autoinhibition is released and is followed by accelerative allosteric feedback autoactivation. Here, using mutagenesis-based kinetic and pulldown analyses, we show that Noonan syndrome Ras mutants I24N, T50I, V152G, and D153V deregulate the autoactivation of SOS to populate their active form. This previously unknown process has been linked so far only to the development of Noonan syndrome. In contrast, other Noonan syndrome Ras mutants-V14I, T58I, and G60E-populate their active form by deregulation of the previously documented Ras GTPase activities. We propose a novel mechanism responsible for the deregulation of SOS autoactivation, where I24N, T50I, V152G, and D153V Ras mutants evade SOS autoinhibition. Consequently, they are capable of forming a complex with the SOS allosteric site, thus aberrantly promoting SOS autoactivation, resulting in the population of active Ras mutants in cells. The results of this study elucidate the molecular mechanism of the Ras mutant-mediated development of Noonan syndrome.
Keywords: Noonan syndrome; Ras; Ras protein; SOS; Son of Sevenless; allosteric regulation; allostery; autoactivation; autoinhibition; catalysis; enzyme kinetics; guanine nucleotide exchange factor (GEF).
© 2020 Umutesi et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures
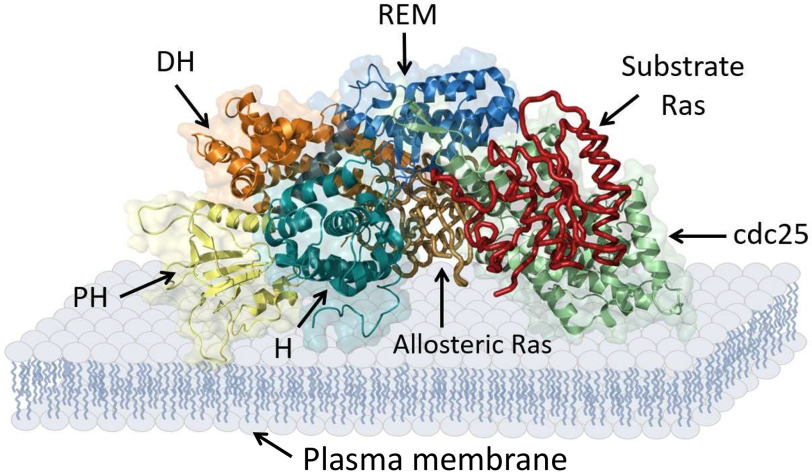
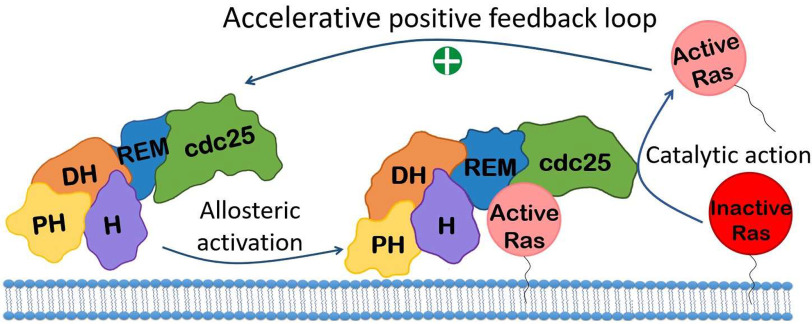
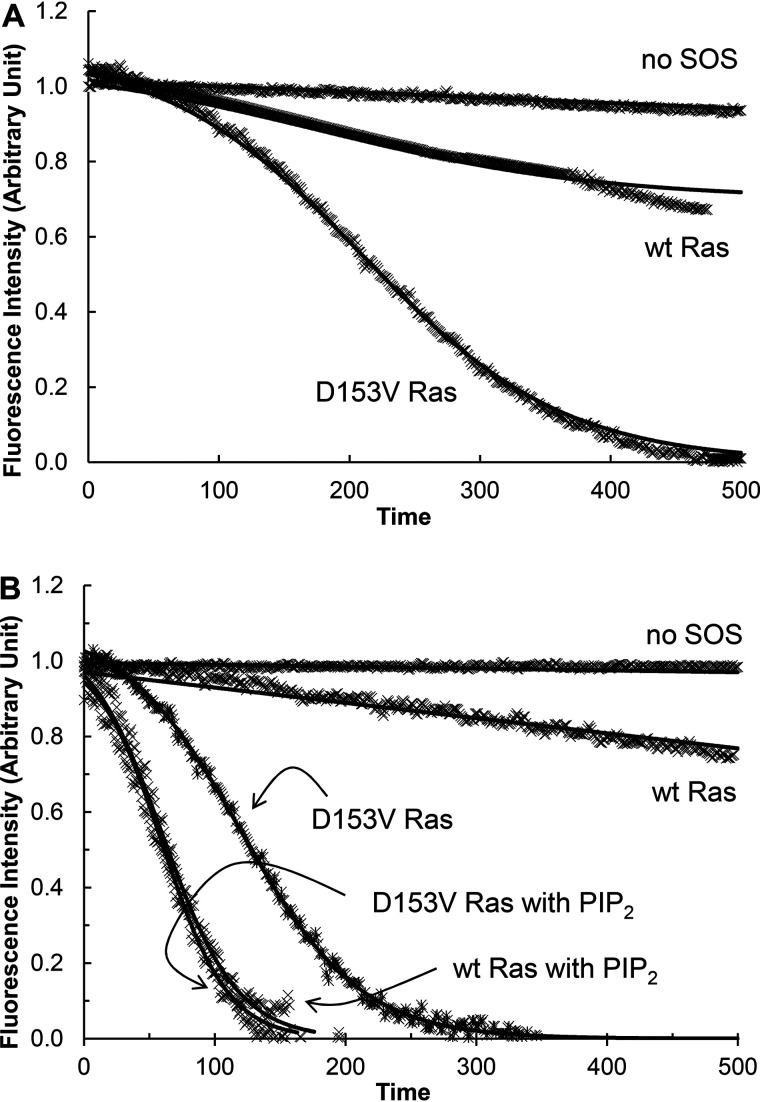
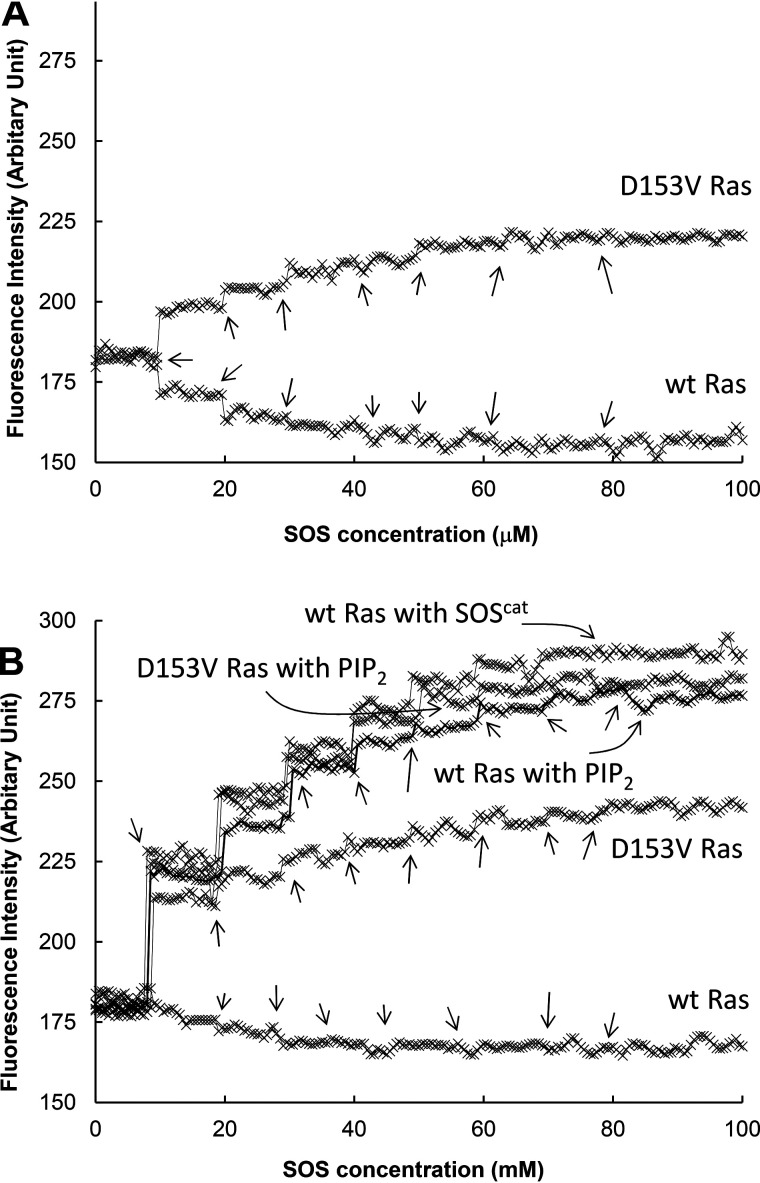
References
-
- Bos J. L. (1989) ras oncogenes in human cancer: a review. Cancer Res. 49, 4682–4689 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
