

Zika Virus Infection Promotes Local Inflammation, Cell Adhesion Molecule Upregulation, and Leukocyte Recruitment at the Blood-Brain Barrier
- PMID: 32753493
- PMCID: PMC7407083
- DOI: 10.1128/mBio.01183-20
Zika Virus Infection Promotes Local Inflammation, Cell Adhesion Molecule Upregulation, and Leukocyte Recruitment at the Blood-Brain Barrier
Abstract
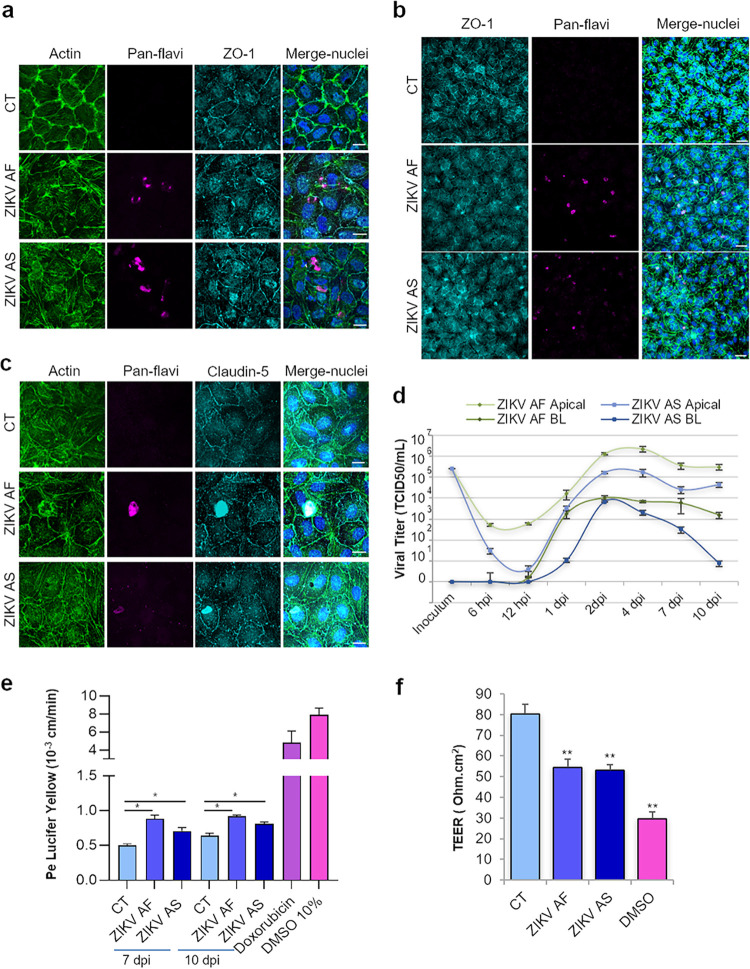
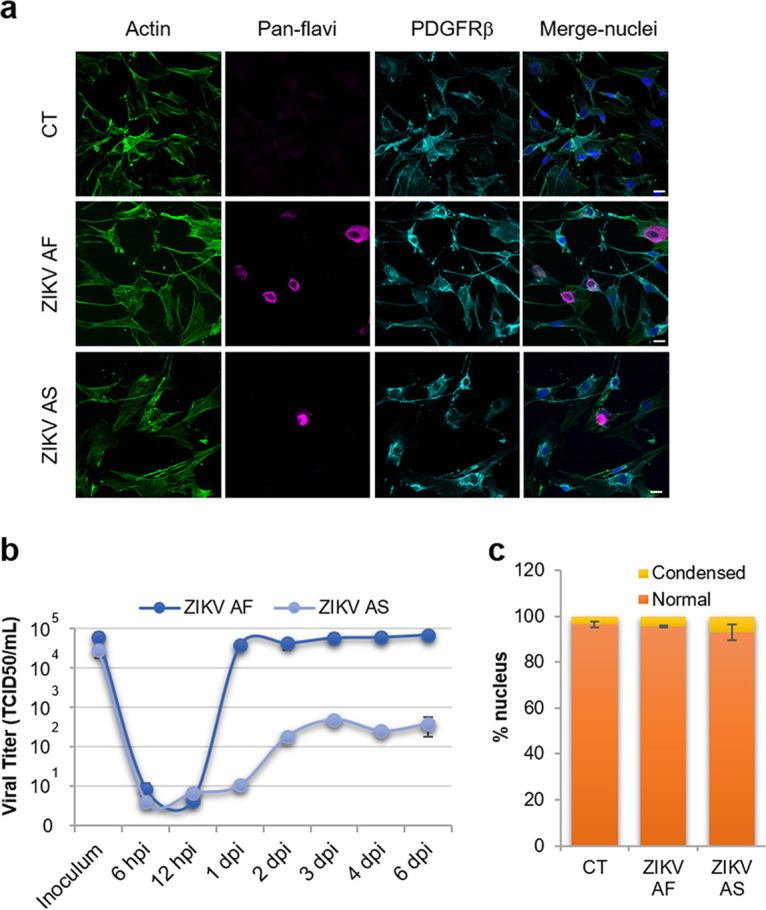
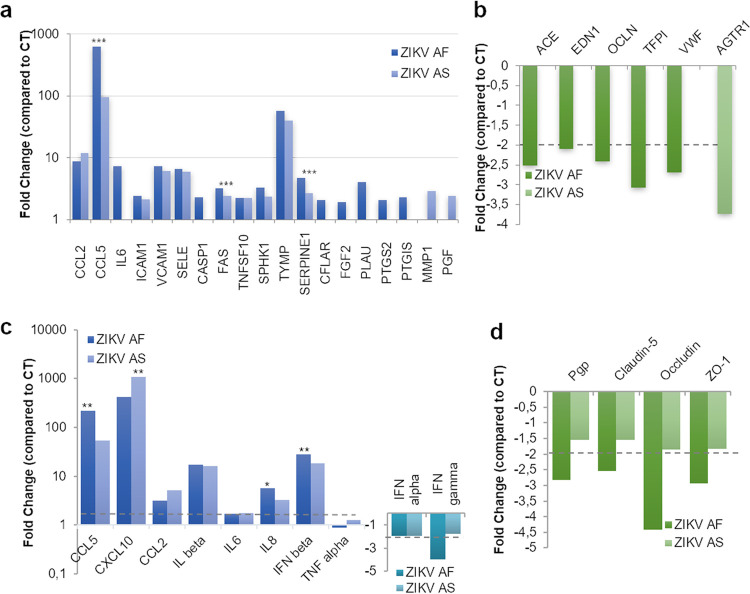
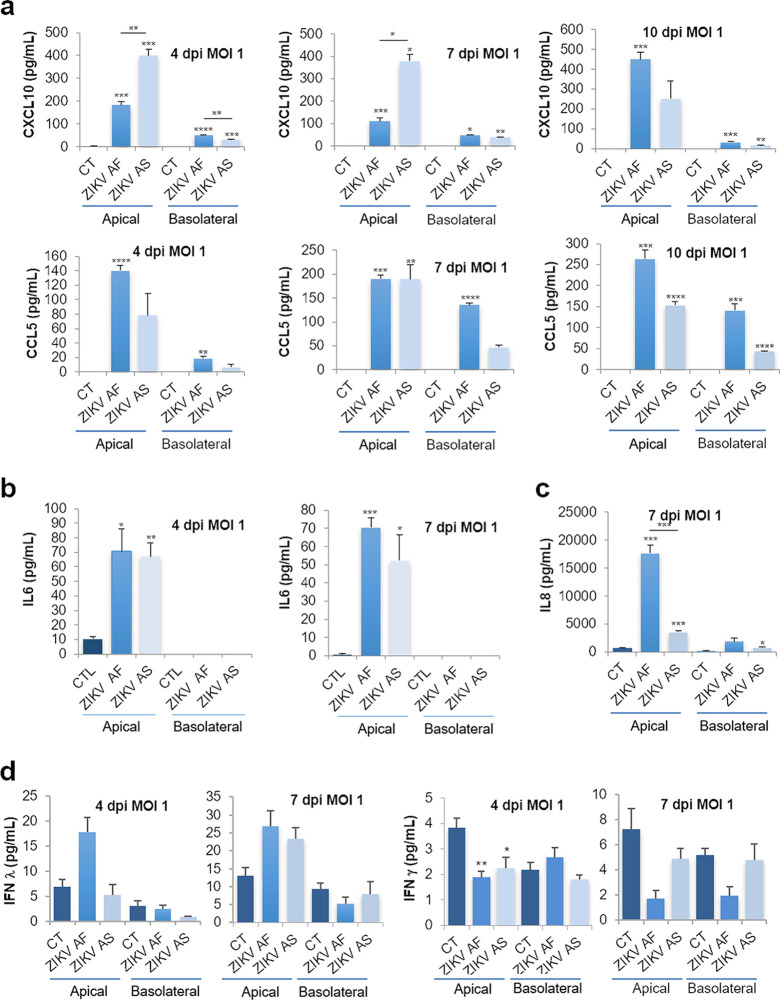
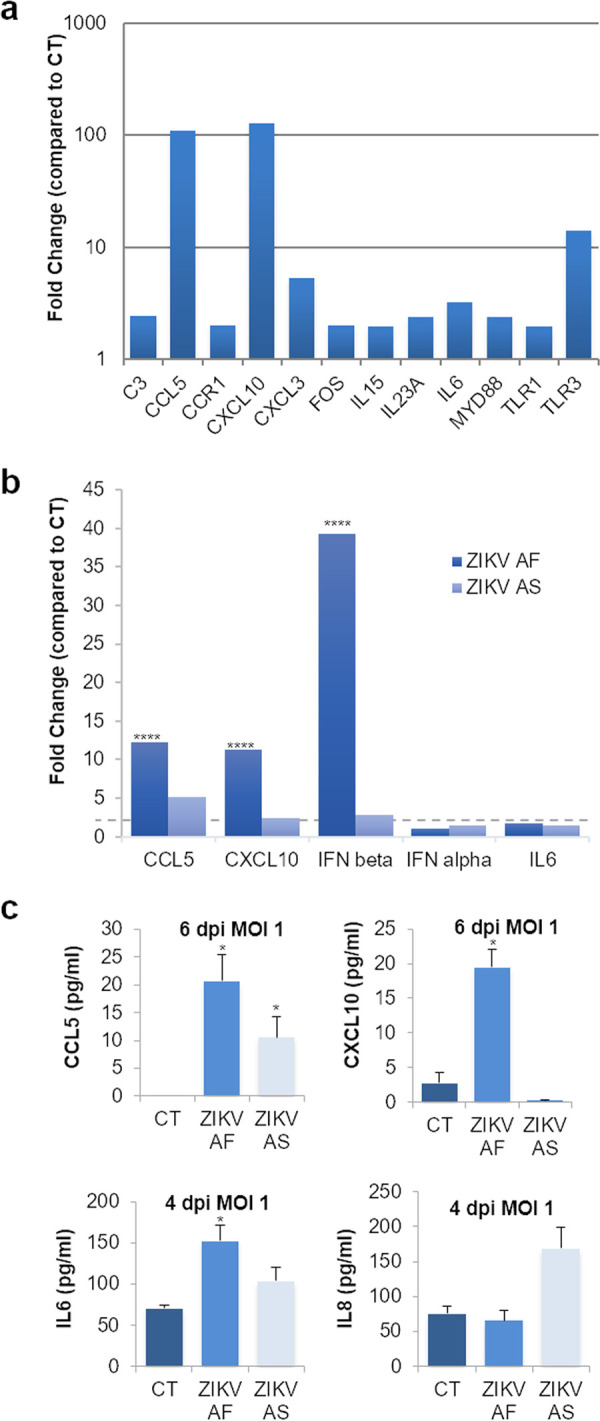
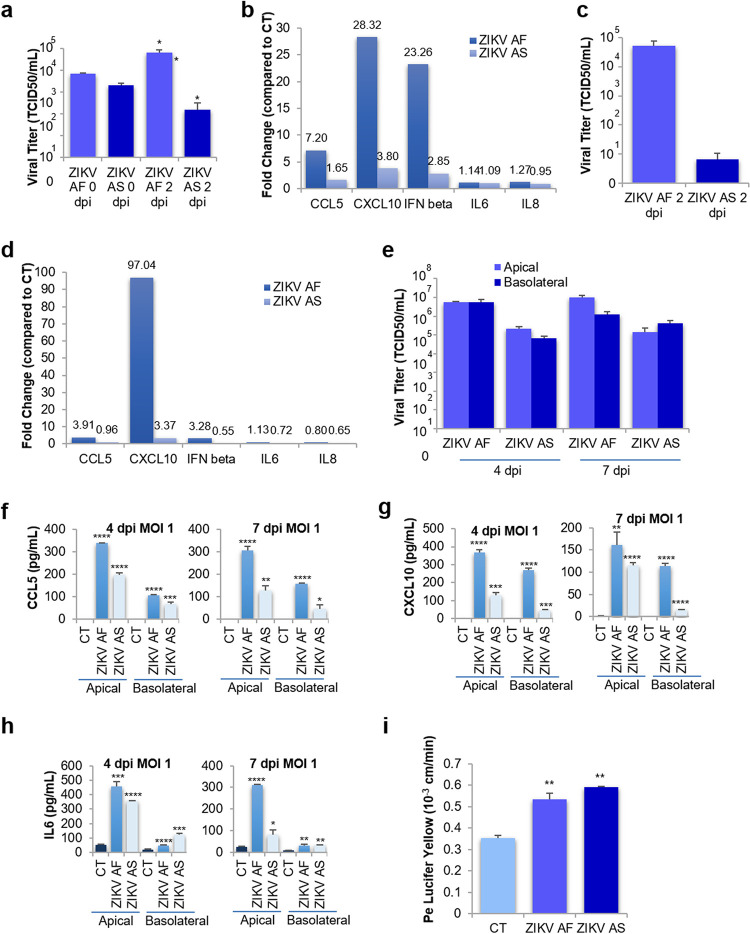
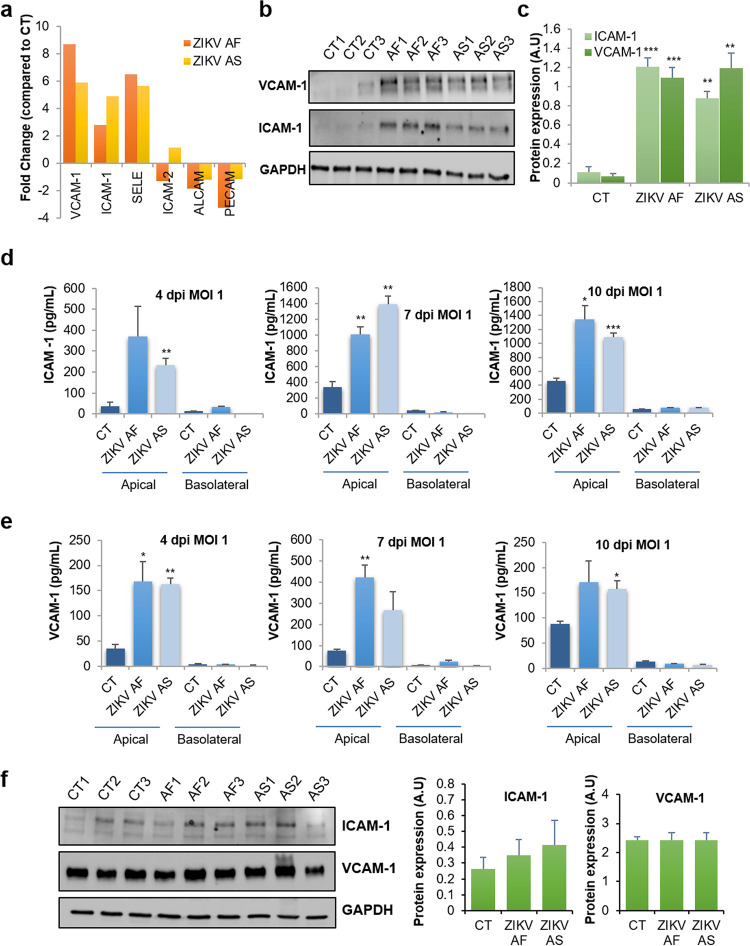
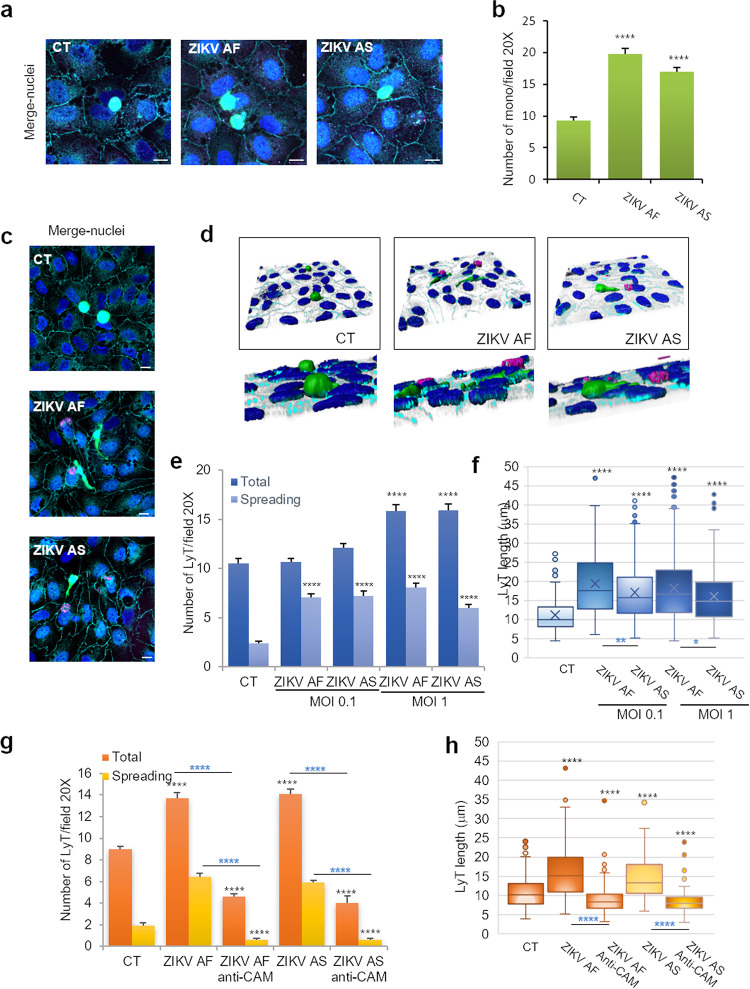
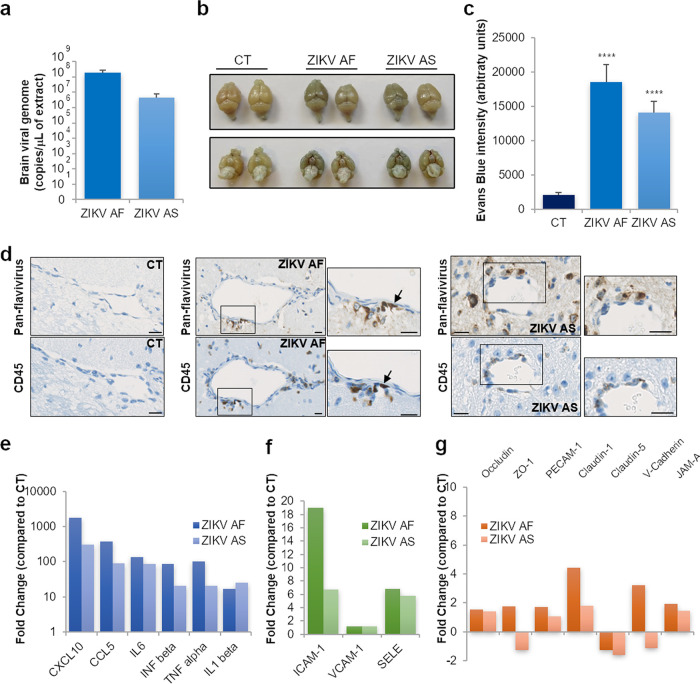
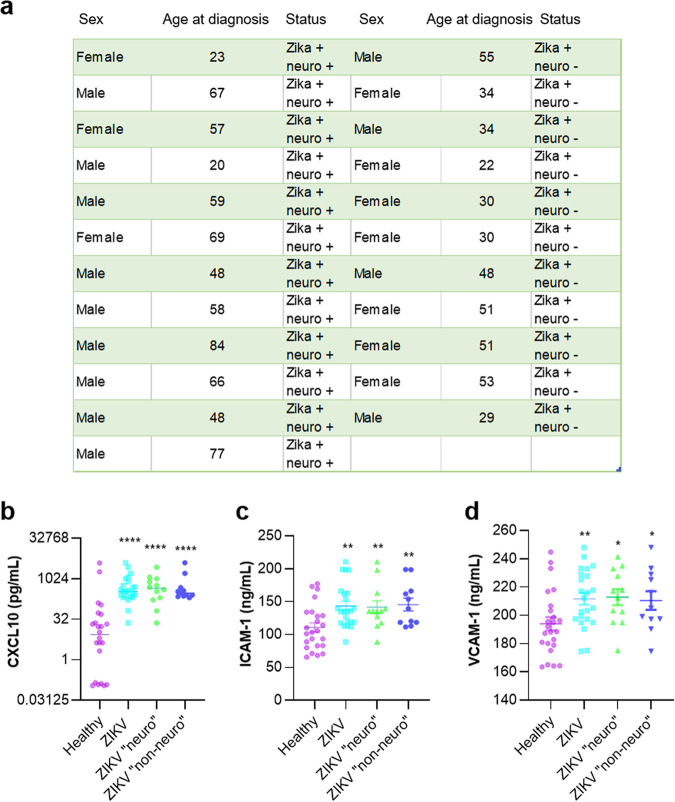
The blood-brain barrier (BBB) largely prevents toxins and pathogens from accessing the brain. Some viruses have the ability to cross this barrier and replicate in the central nervous system (CNS). Zika virus (ZIKV) was responsible in 2015 to 2016 for a major epidemic in South America and was associated in some cases with neurological impairments. Here, we characterized some of the mechanisms behind its neuroinvasion using an innovative in vitro human BBB model. ZIKV efficiently replicated, was released on the BBB parenchyma side, and triggered subtle modulation of BBB integrity as well as an upregulation of inflammatory and cell adhesion molecules (CAMs), which in turn favored leukocyte recruitment. Finally, we showed that ZIKV-infected mouse models displayed similar CAM upregulation and that soluble CAMs were increased in plasma samples from ZIKV-infected patients. Our observations suggest a complex interplay between ZIKV and the BBB, which may trigger local inflammation, leukocyte recruitment, and possible cerebral vasculature impairment.IMPORTANCE Zika virus (ZIKV) can be associated with neurological impairment in children and adults. To reach the central nervous system, viruses have to cross the blood-brain barrier (BBB), a multicellular system allowing a tight separation between the bloodstream and the brain. Here, we show that ZIKV infects cells of the BBB and triggers a subtle change in its permeability. Moreover, ZIKV infection leads to the production of inflammatory molecules known to modulate BBB integrity and participate in immune cell attraction. The virus also led to the upregulation of cellular adhesion molecules (CAMs), which in turn favored immune cell binding to the BBB and potentially increased infiltration into the brain. These results were also observed in a mouse model of ZIKV infection. Furthermore, plasma samples from ZIKV-infected patients displayed an increase in CAMs, suggesting that this mechanism could be involved in neuroinflammation triggered by ZIKV.
Keywords: Zika virus; blood-brain barrier; cell adhesion molecules; leukocyte recruitment.
Copyright © 2020 Clé et al.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
