

Lipo-chitooligosaccharides as regulatory signals of fungal growth and development
- PMID: 32753587
- PMCID: PMC7403392
- DOI: 10.1038/s41467-020-17615-5
Lipo-chitooligosaccharides as regulatory signals of fungal growth and development
Abstract
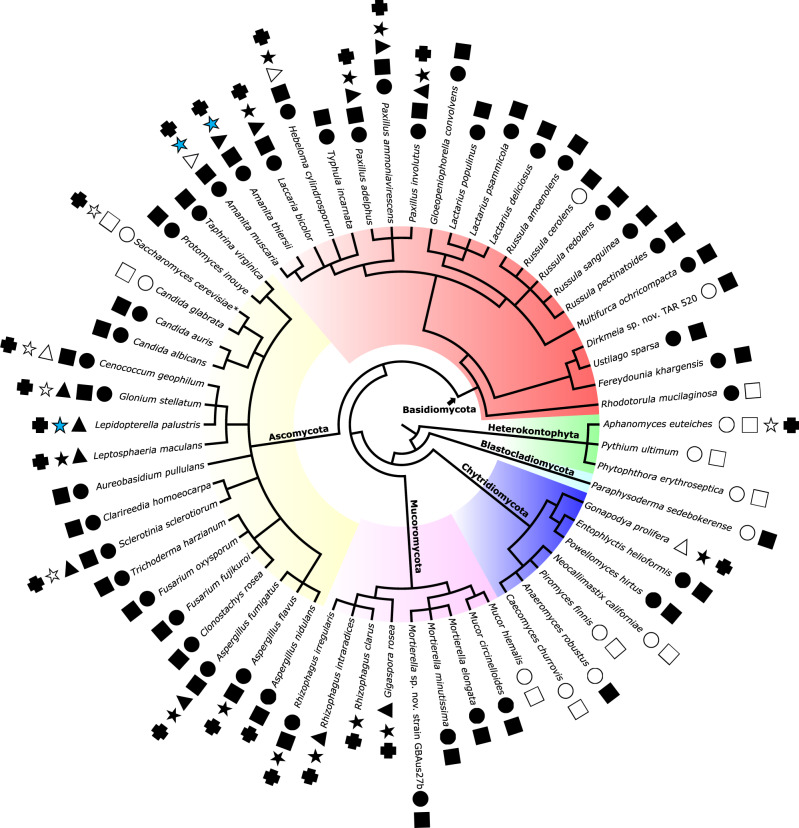
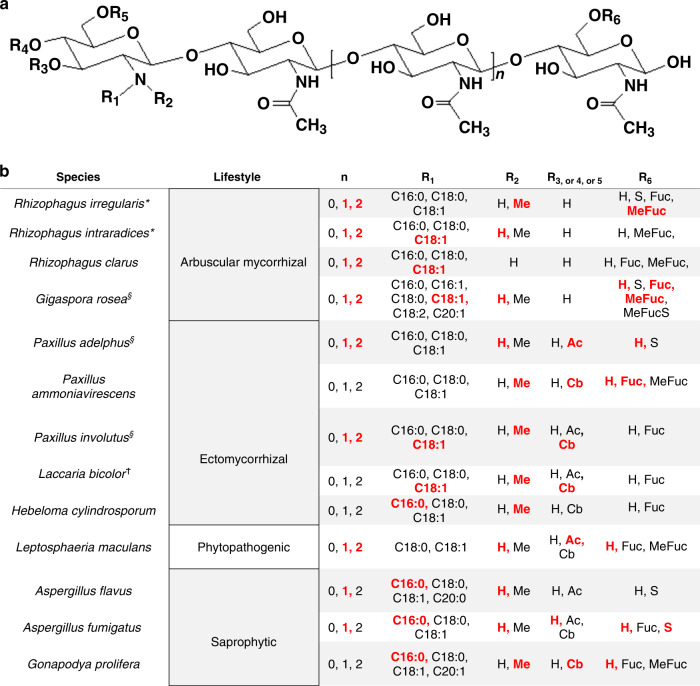
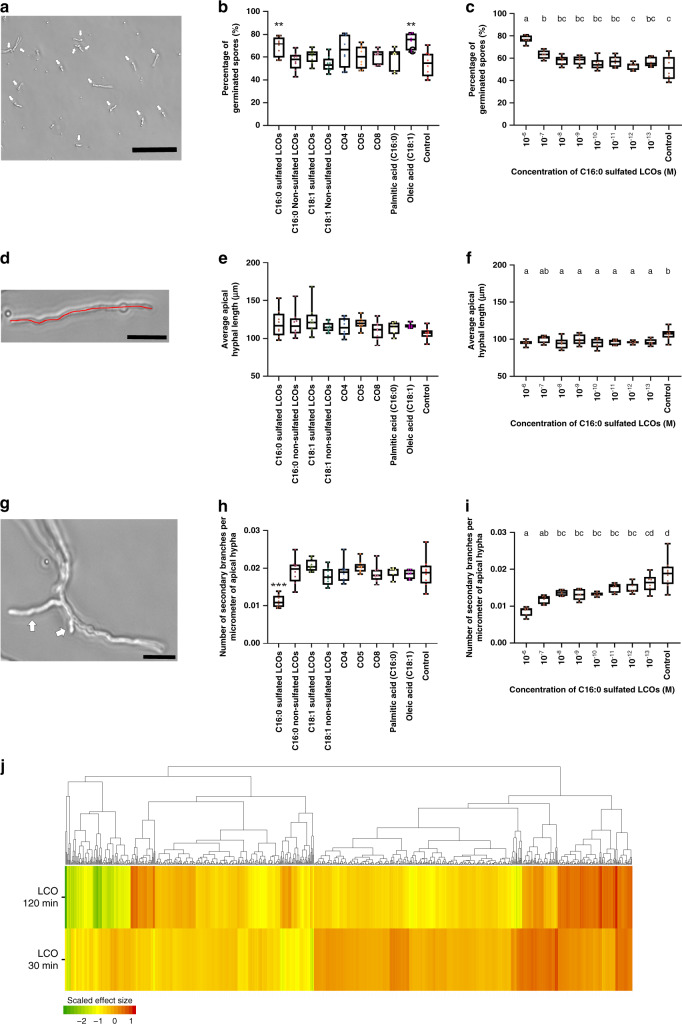
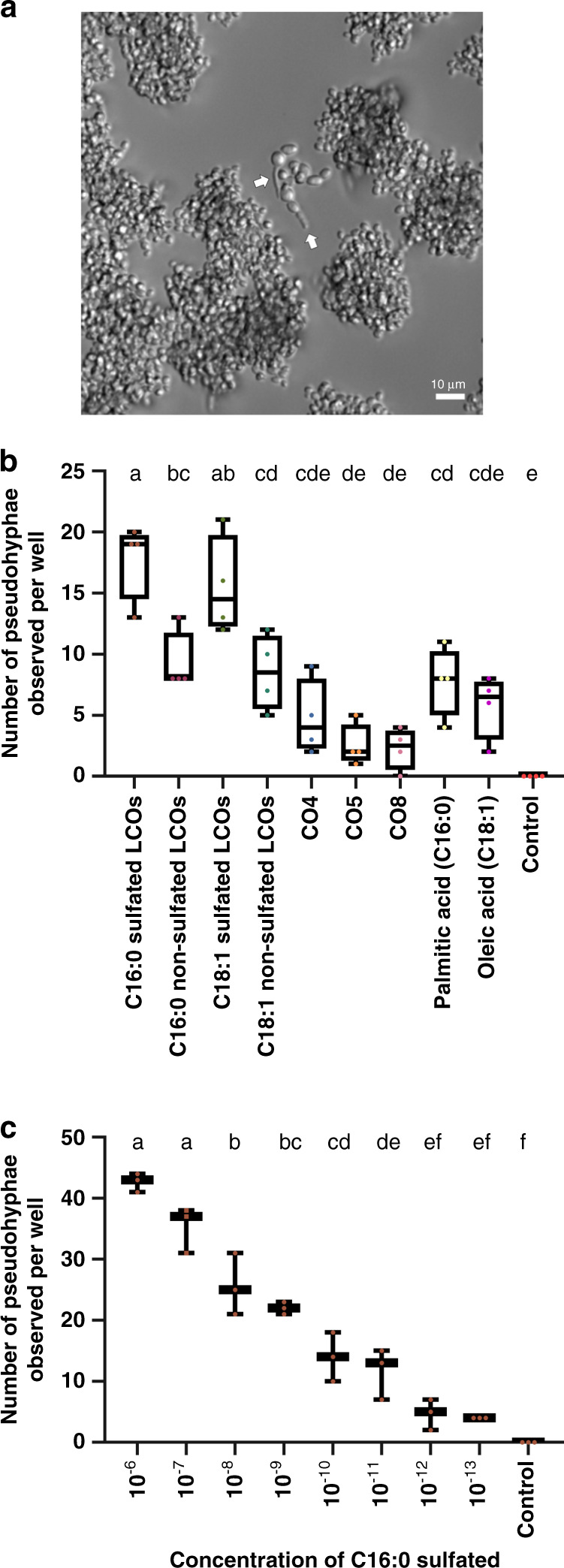
Lipo-chitooligosaccharides (LCOs) are signaling molecules produced by rhizobial bacteria that trigger the nodulation process in legumes, and by some fungi that also establish symbiotic relationships with plants, notably the arbuscular and ecto mycorrhizal fungi. Here, we show that many other fungi also produce LCOs. We tested 59 species representing most fungal phyla, and found that 53 species produce LCOs that can be detected by functional assays and/or by mass spectroscopy. LCO treatment affects spore germination, branching of hyphae, pseudohyphal growth, and transcription in non-symbiotic fungi from the Ascomycete and Basidiomycete phyla. Our findings suggest that LCO production is common among fungi, and LCOs may function as signals regulating fungal growth and development.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Lerouge P, et al. Symbiotic host-specificity of Rhizobium meliloti is determined by a sulphated and acylated glucosamine oligosaccharide signal. Nature. 1990;344:781–784. - PubMed
-
- Dénarié J, Debellé F, Promé J-C. Rhizobium lipo-chitooligosaccharide nodulation factors: signaling molecules mediating recognition and morphogenesis. Annu. Rev. Biochem. 1996;65:503–535. - PubMed
-
- Maillet F, et al. Fungal lipochitooligosaccharide symbiotic signals in arbuscular mycorrhiza. Nature. 2011;469:58–63. - PubMed
-
- Venkateshwaran M, Volkening JD, Sussman MR, Ané J-M. Symbiosis and the social network of higher plants. Curr. Opin. Plant Biol. 2013;16:118–127. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
