

Heme Synthesis Inhibition Blocks Angiogenesis via Mitochondrial Dysfunction
- PMID: 32755804
- PMCID: PMC7399258
- DOI: 10.1016/j.isci.2020.101391
Heme Synthesis Inhibition Blocks Angiogenesis via Mitochondrial Dysfunction
Abstract
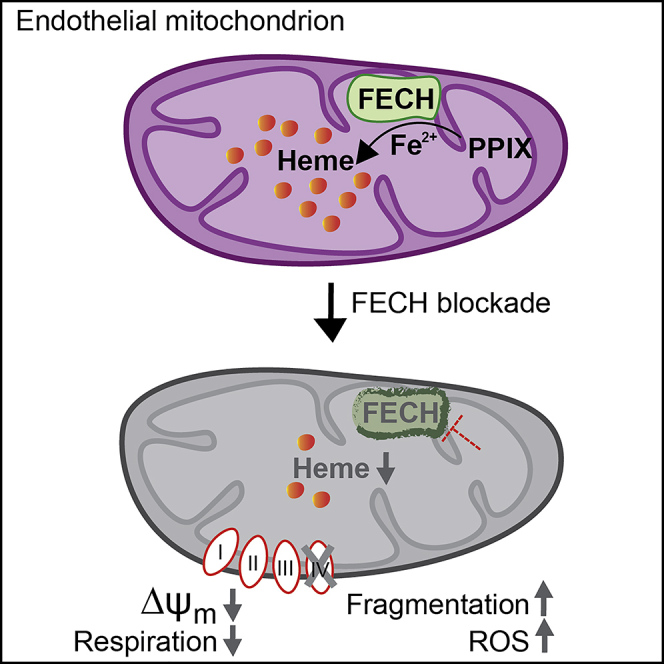
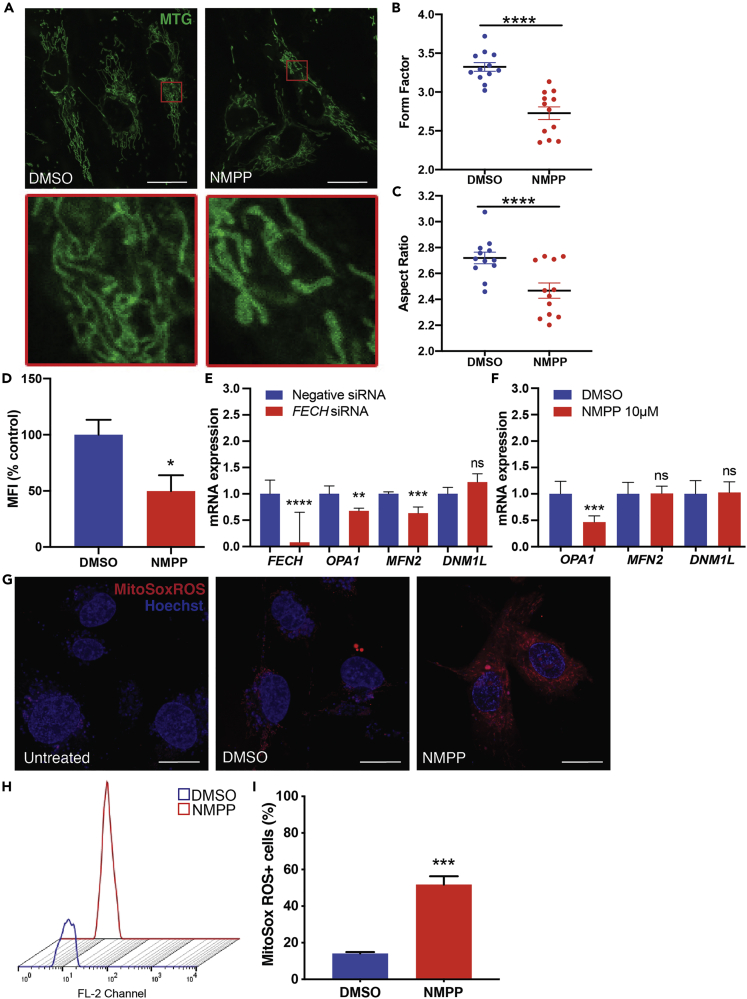
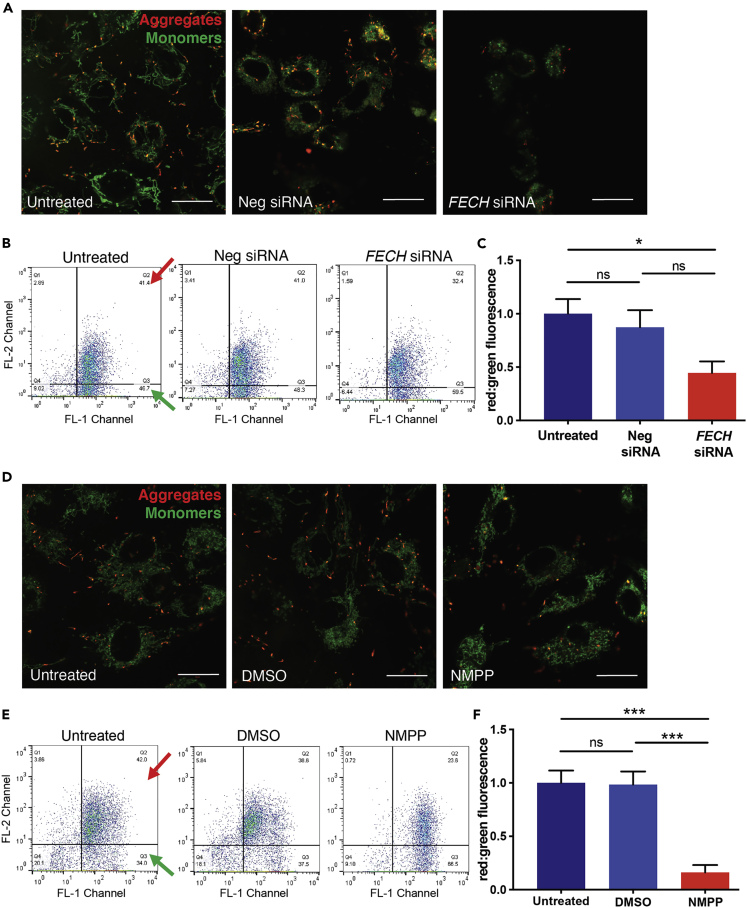
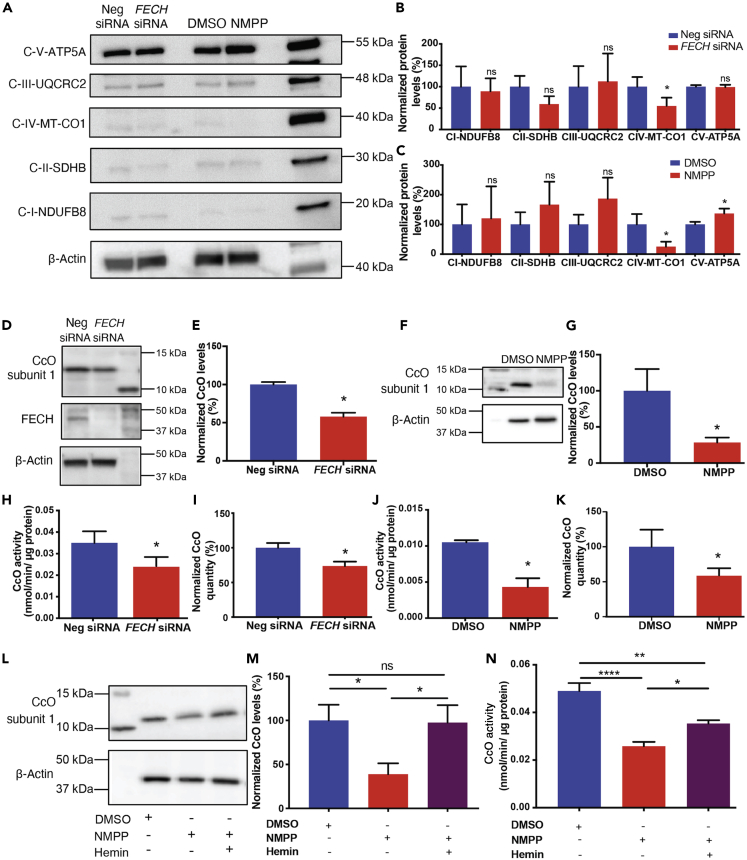
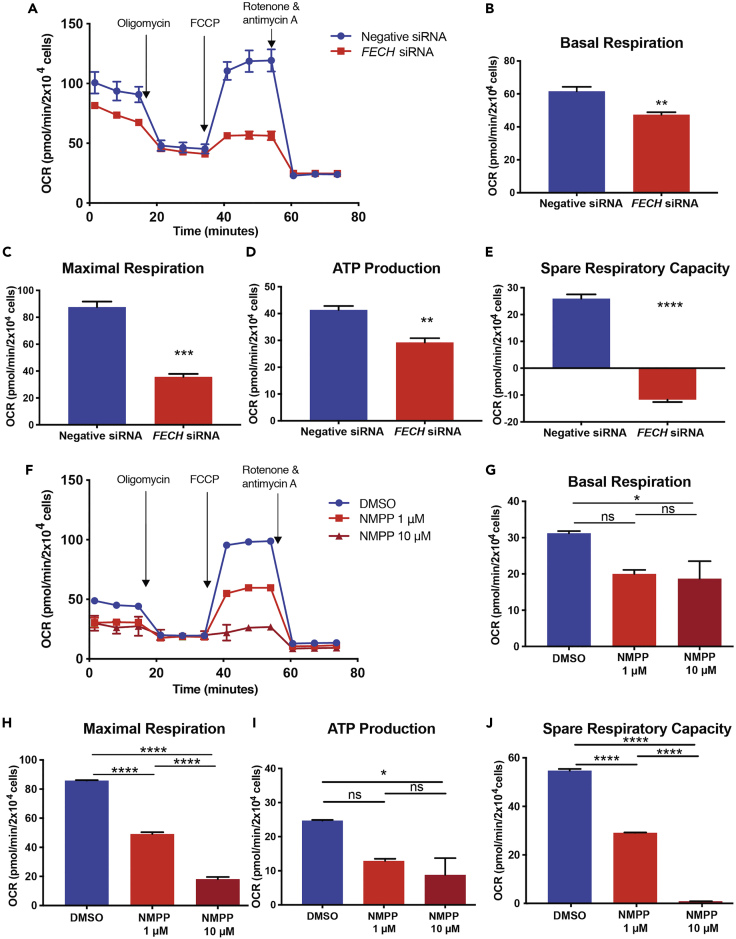
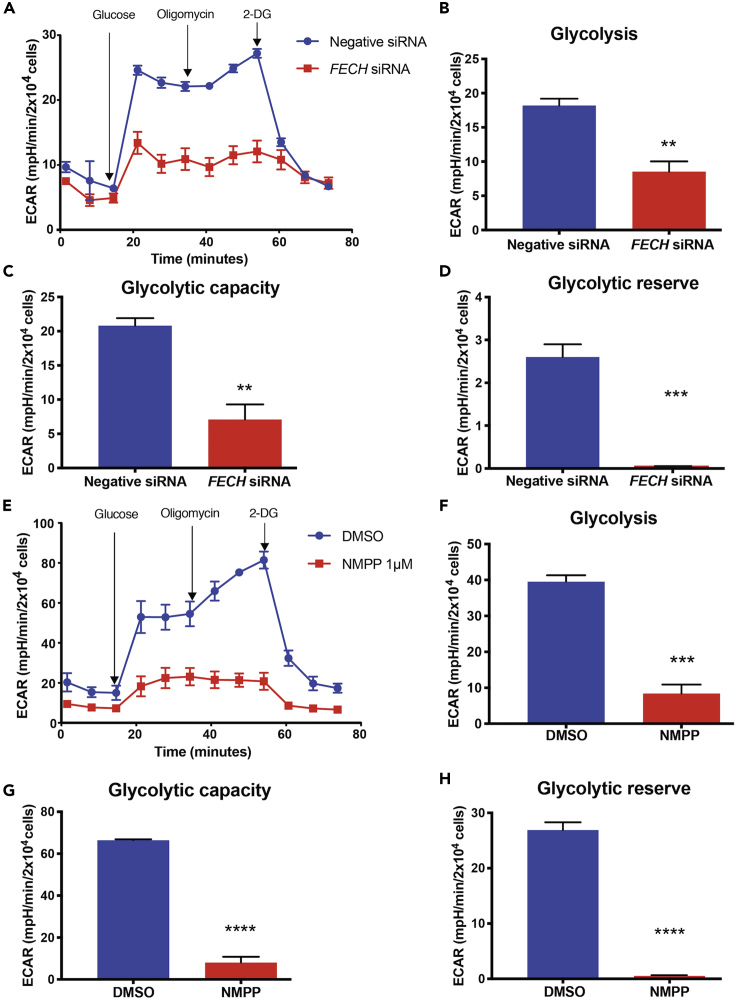
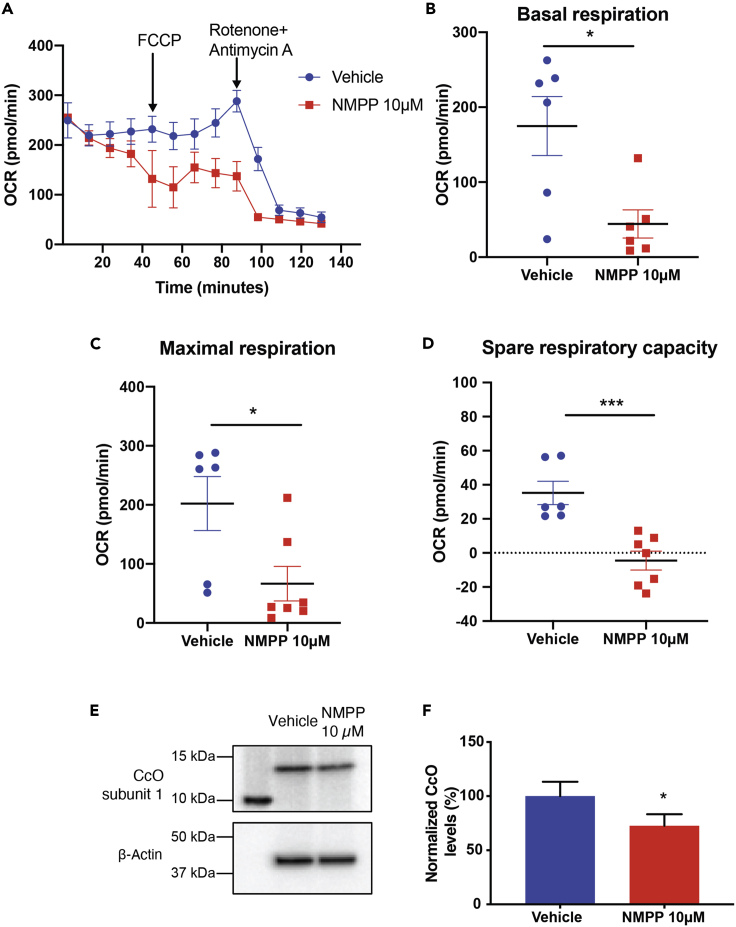
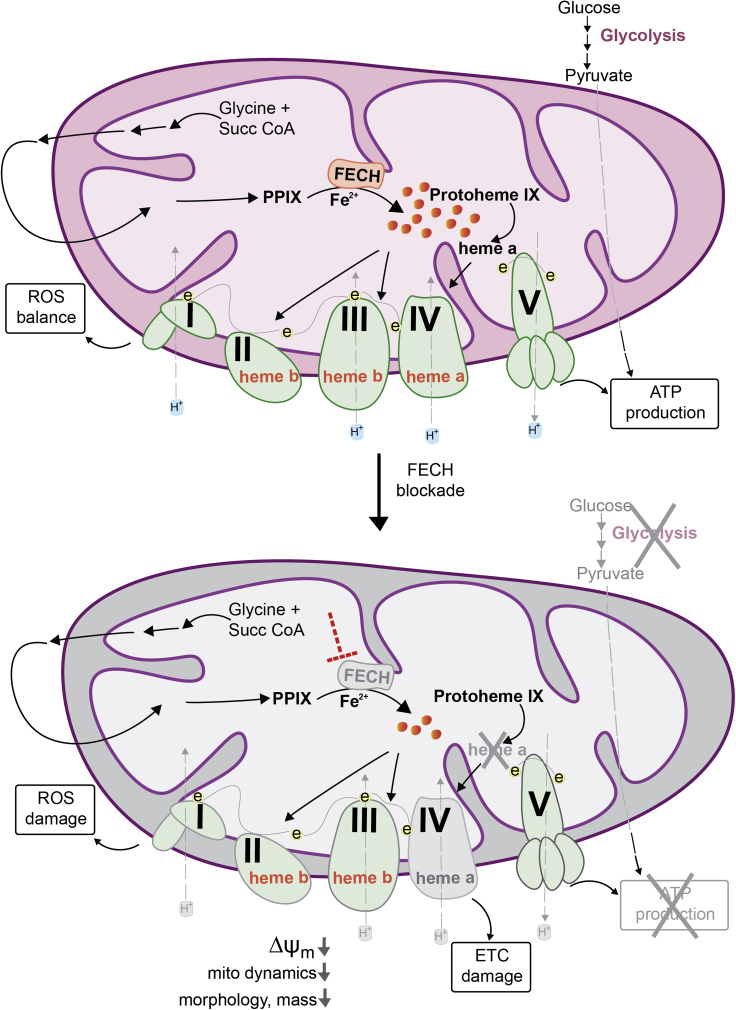
The relationship between heme metabolism and angiogenesis is poorly understood. The final synthesis of heme occurs in mitochondria, where ferrochelatase (FECH) inserts Fe2+ into protoporphyrin IX to produce proto-heme IX. We previously showed that FECH inhibition is antiangiogenic in human retinal microvascular endothelial cells (HRECs) and in animal models of ocular neovascularization. In the present study, we sought to understand the mechanism of how FECH and thus heme is involved in endothelial cell function. Mitochondria in endothelial cells had several defects in function after heme inhibition. FECH loss changed the shape and mass of mitochondria and led to significant oxidative stress. Oxidative phosphorylation and mitochondrial Complex IV were decreased in HRECs and in murine retina ex vivo after heme depletion. Supplementation with heme partially rescued phenotypes of FECH blockade. These findings provide an unexpected link between mitochondrial heme metabolism and angiogenesis.
Keywords: Cell Biology; Developmental Genetics; Physiology.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests T.W.C. is a named inventor on patent applications related to this work. The other authors declare no competing interests.
Figures
References
-
- Acin-Perez R., Fernandez-Silva P., Peleato M.L., Perez-Martos A., Enriquez J.A. Respiratory active mitochondrial supercomplexes. Mol. Cell. 2008;32:529–539. - PubMed
-
- Atamna H., Liu J., Ames B.N. Heme deficiency selectively interrupts assembly of mitochondrial complex IV in human fibroblasts: revelance to aging. J. Biol. Chem. 2001;276:48410–48416. - PubMed
-
- De Bock K., Georgiadou M., Schoors S., Kuchnio A., Wong B.W., Cantelmo A.R., Quaegebeur A., Ghesquiere B., Cauwenberghs S., Eelen G. Role of PFKFB3-driven glycolysis in vessel sprouting. Cell. 2013;154:651–663. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
