

Induction of a Timed Metabolic Collapse to Overcome Cancer Chemoresistance
- PMID: 32763164
- PMCID: PMC8397232
- DOI: 10.1016/j.cmet.2020.07.009
Induction of a Timed Metabolic Collapse to Overcome Cancer Chemoresistance
Abstract
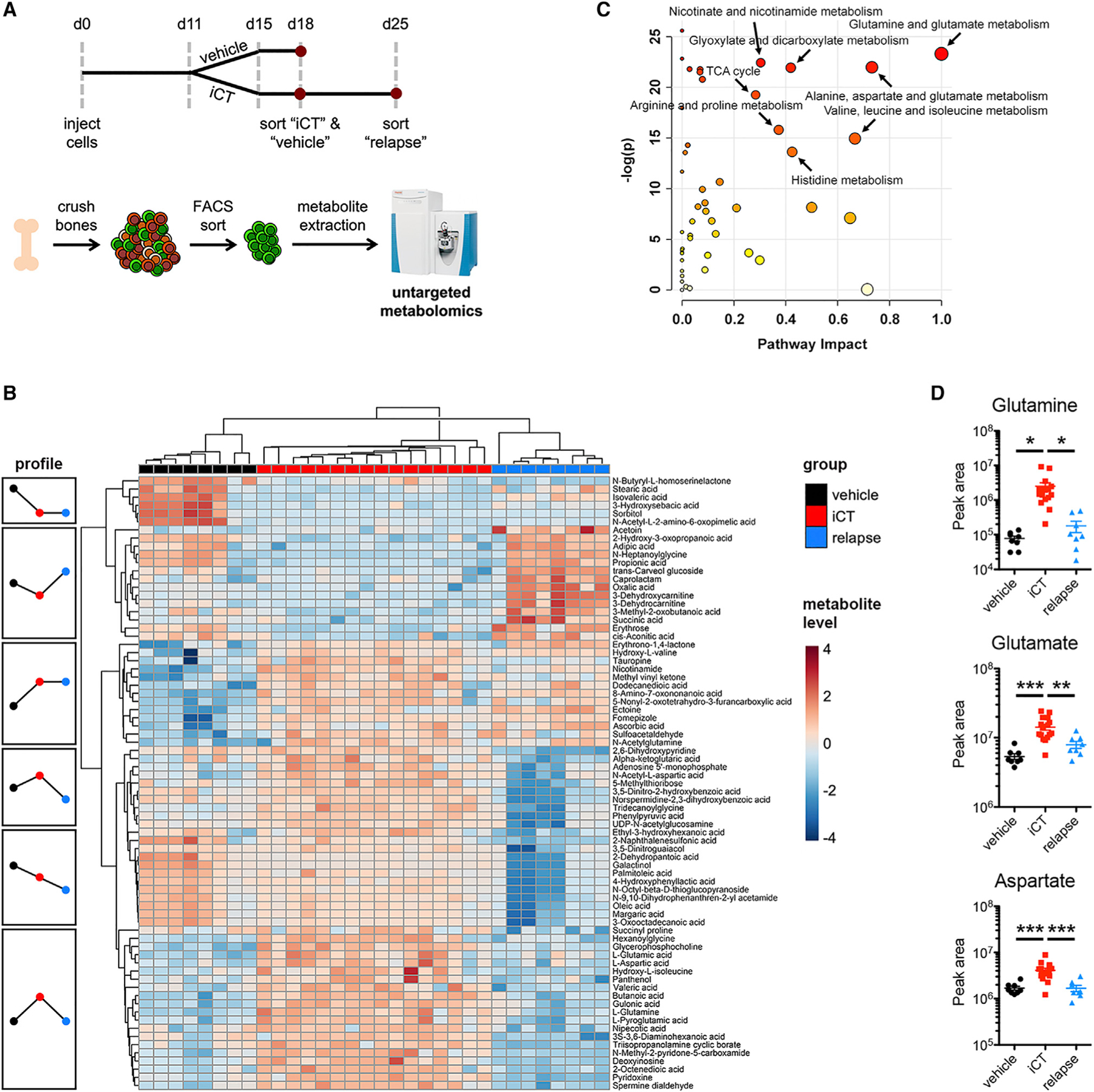
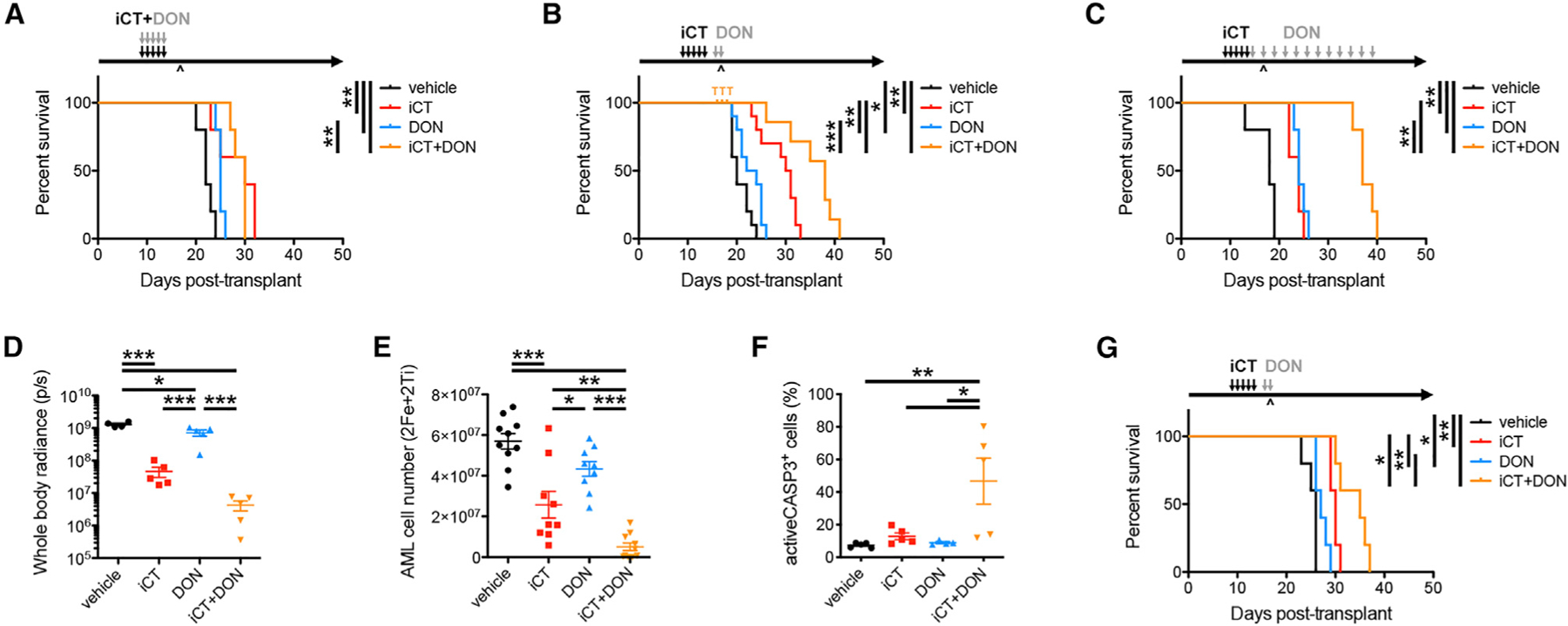
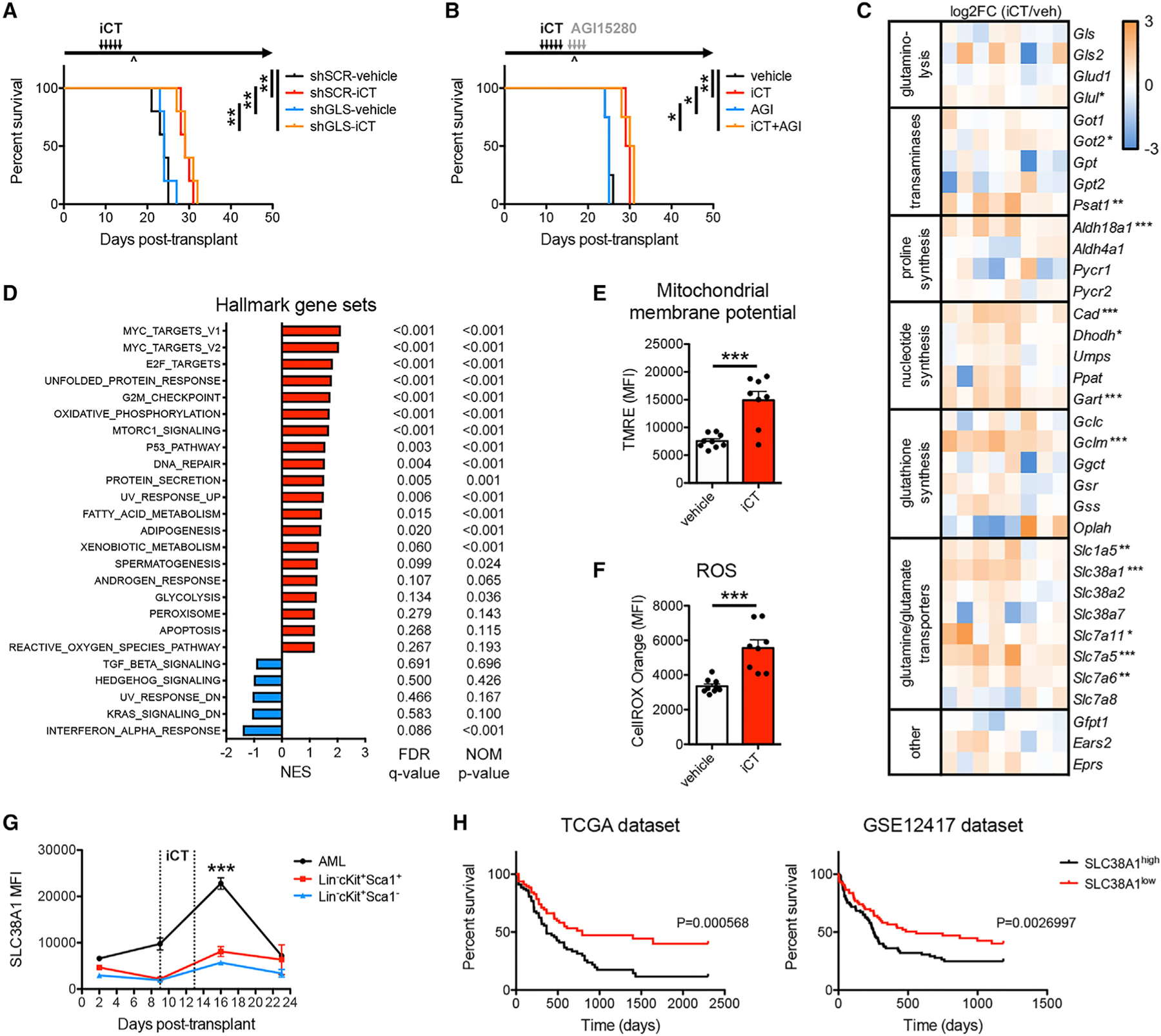
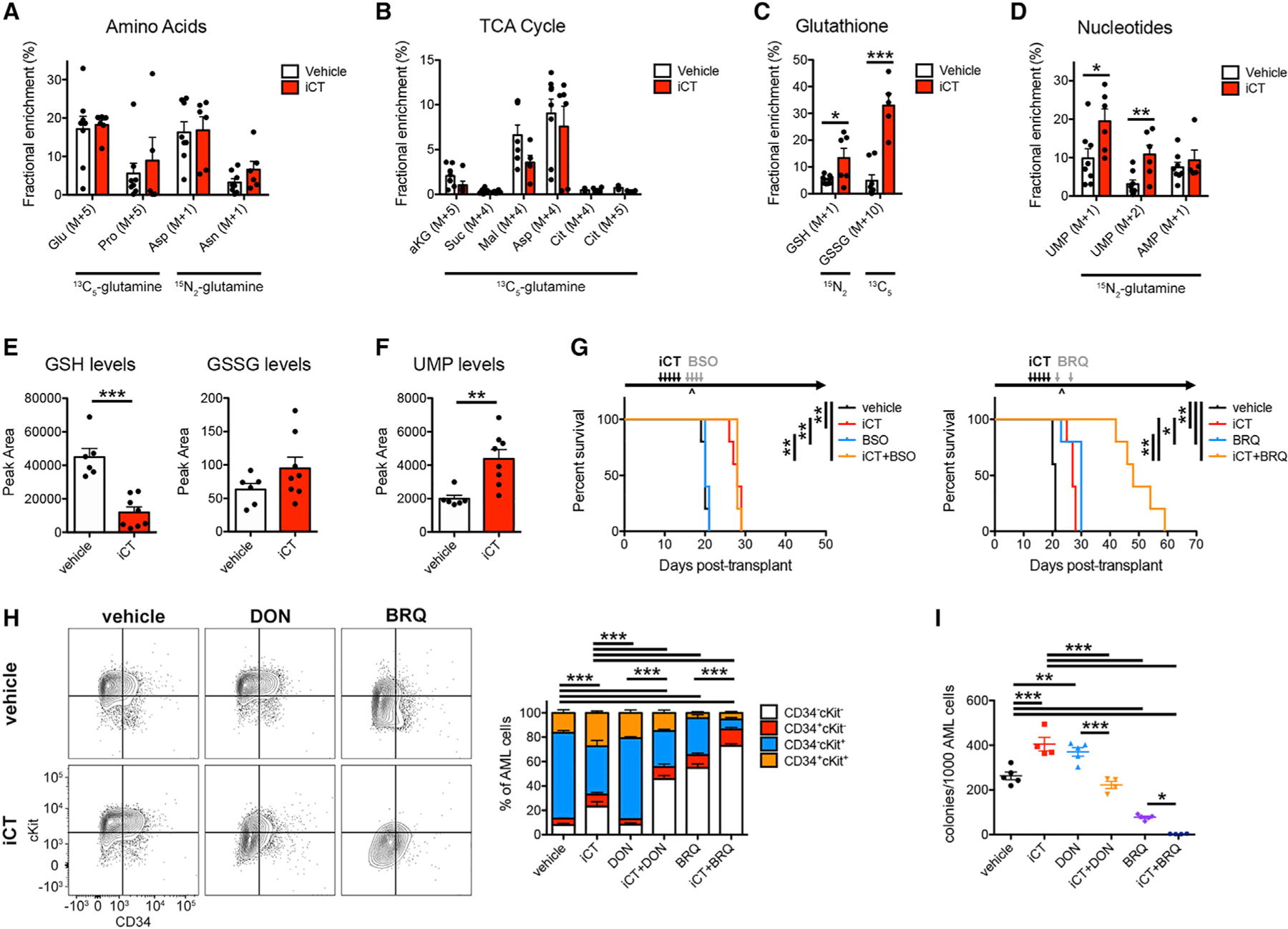
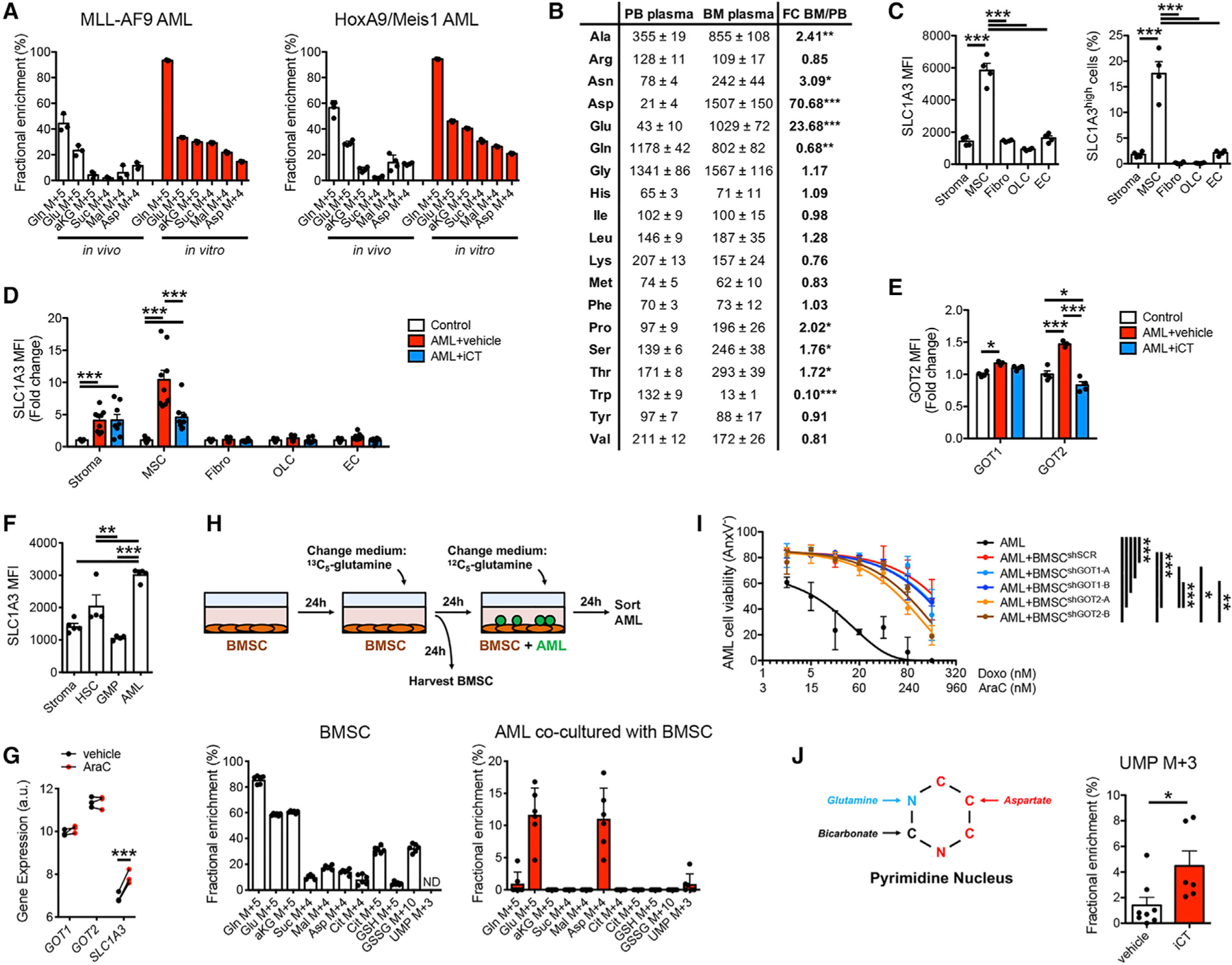
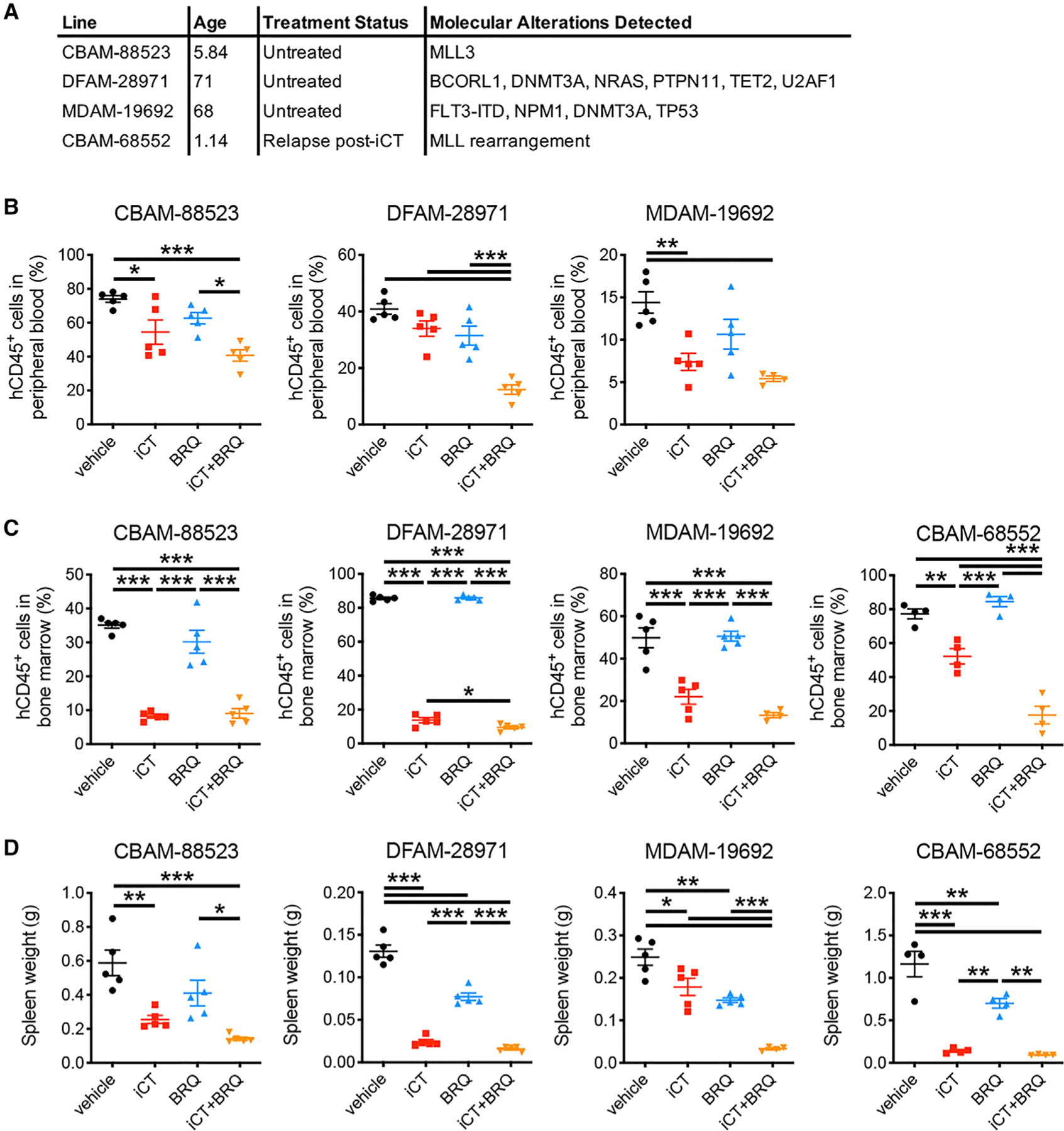
Cancer relapse begins when malignant cells pass through the extreme metabolic bottleneck of stress from chemotherapy and the byproducts of the massive cell death in the surrounding region. In acute myeloid leukemia, complete remissions are common, but few are cured. We tracked leukemia cells in vivo, defined the moment of maximal response following chemotherapy, captured persisting cells, and conducted unbiased metabolomics, revealing a metabolite profile distinct from the pre-chemo growth or post-chemo relapse phase. Persisting cells used glutamine in a distinctive manner, preferentially fueling pyrimidine and glutathione generation, but not the mitochondrial tricarboxylic acid cycle. Notably, malignant cell pyrimidine synthesis also required aspartate provided by specific bone marrow stromal cells. Blunting glutamine metabolism or pyrimidine synthesis selected against residual leukemia-initiating cells and improved survival in leukemia mouse models and patient-derived xenografts. We propose that timed cell-intrinsic or niche-focused metabolic disruption can exploit a transient vulnerability and induce metabolic collapse in cancer cells to overcome chemoresistance.
Keywords: acute myeloid leukemia; aspartate; bone marrow niche; cell metabolism; chemotherapy; glutamine; mouse models; patient-derived xenografts; pyrimidine synthesis; tumor microenvironment.
Copyright © 2020. Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interests D.B.S. is co-founder and owns equity in Clear Creek Bio. P.P.H. is a consultant for Auron Therapeutics. M.G.V.H. is a consultant and SAB member for Agios Pharmaceuticals, Aeglea Biotherapeutics, iTeos Therapeutics, and Auron Therapeutics. D.T.S. is a director and equity holder of Agios Pharmaceuticals, Magenta Therapeutics, Editas Medicines, Clear Creek Bio, and LifeVaultBio; he is a founder of Fate Therapeutics and Magenta Therapeutics and a consultant to FOG Pharma, VcanBio, and Flagship Pioneering. N.v.G., A. Schajnovitz, T.O., and D.T.S. are inventors on patents related to this work.
Figures
Comment in
-
Microenvironmental Aspartate Preserves Leukemic Cells from Therapy-Induced Metabolic Collapse.Cell Metab. 2020 Sep 1;32(3):321-323. doi: 10.1016/j.cmet.2020.08.008. Cell Metab. 2020. PMID: 32877685
References
-
- Bajzikova M, Kovarova J, Coelho AR, Boukalova S, Oh S, Rohlenova K, Svec D, Hubackova S, Endaya B, Judasova K, et al. (2019). Reactivation of dihydroorotate dehydrogenase-driven pyrimidine biosynthesis restores tumor growth of respiration-deficient cancer cells. Cell Metab 29, 399–416.e10. - PMC - PubMed
-
- Boukalova S, Hubackova S, Milosevic M, Ezrova Z, Neuzil J, and Rohlena J (2020). Dihydroorotate dehydrogenase in oxidative phosphorylation and cancer. Biochim. Biophys. Acta Mol. Basis Dis. 1866, 165759. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
