

Apolipoprotein L1-Specific Antibodies Detect Endogenous APOL1 inside the Endoplasmic Reticulum and on the Plasma Membrane of Podocytes
- PMID: 32764142
- PMCID: PMC7461670
- DOI: 10.1681/ASN.2019080829
Apolipoprotein L1-Specific Antibodies Detect Endogenous APOL1 inside the Endoplasmic Reticulum and on the Plasma Membrane of Podocytes
Abstract
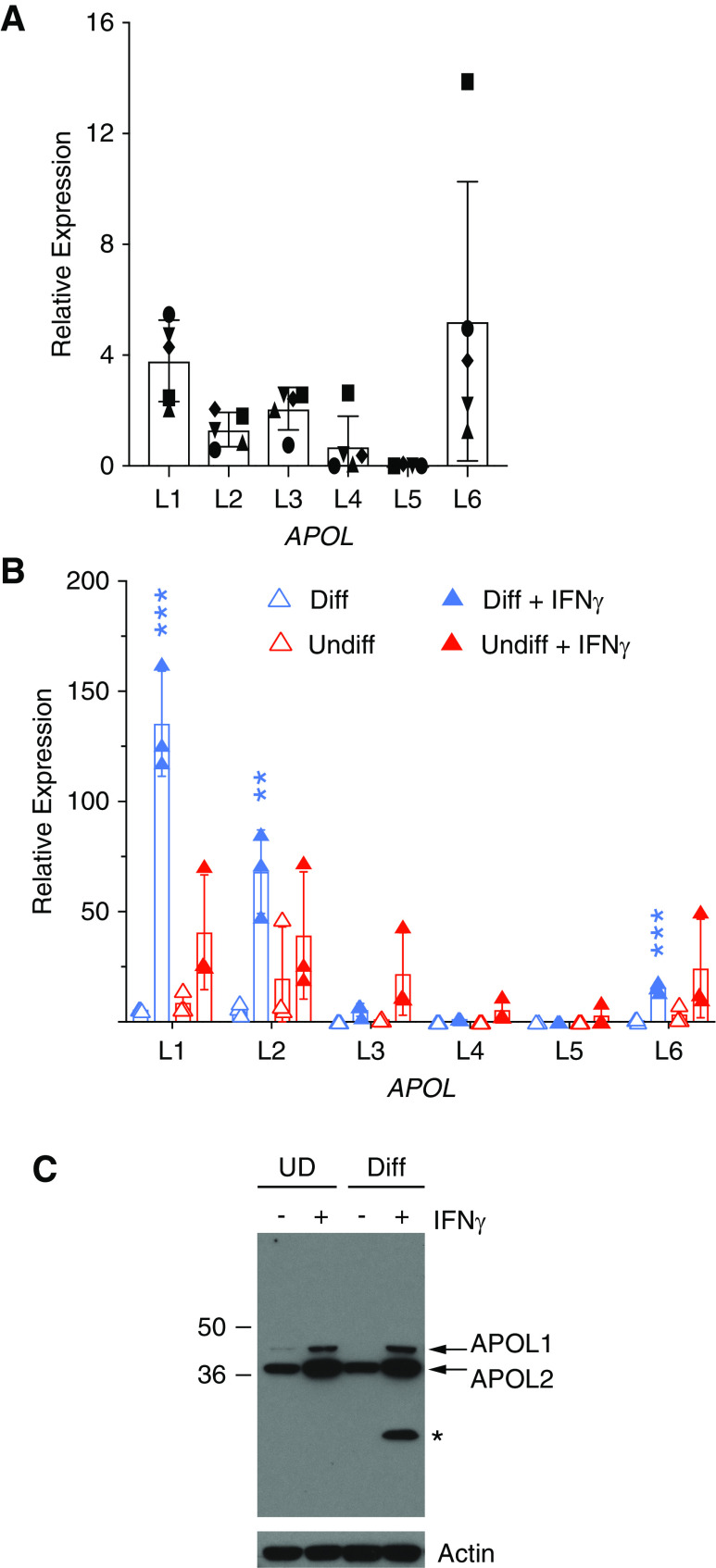
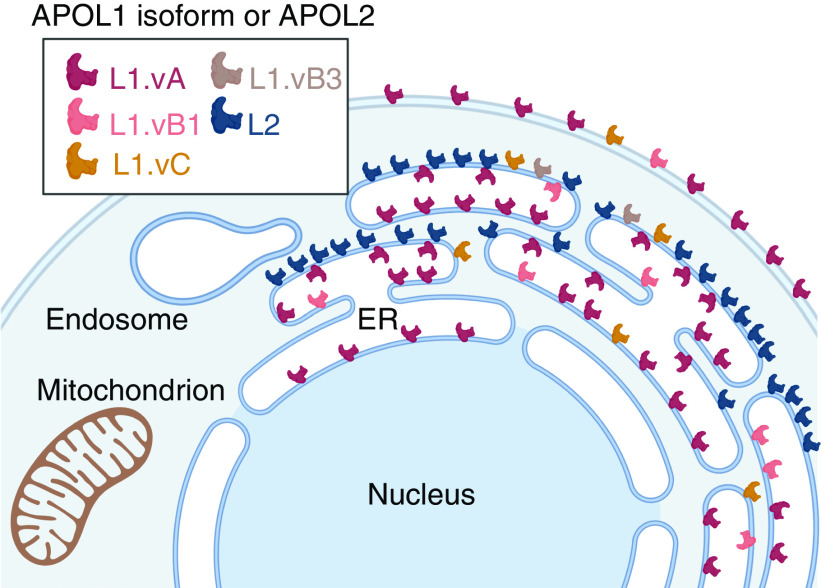
Background: APOL1 is found in human kidney podocytes and endothelia. Variants G1 and G2 of the APOL1 gene account for the high frequency of nondiabetic CKD among African Americans. Proposed mechanisms of kidney podocyte cytotoxicity resulting from APOL1 variant overexpression implicate different subcellular compartments. It is unclear where endogenous podocyte APOL1 resides, because previous immunolocalization studies utilized overexpressed protein or commercially available antibodies that crossreact with APOL2. This study describes and distinguishes the locations of both APOLs.
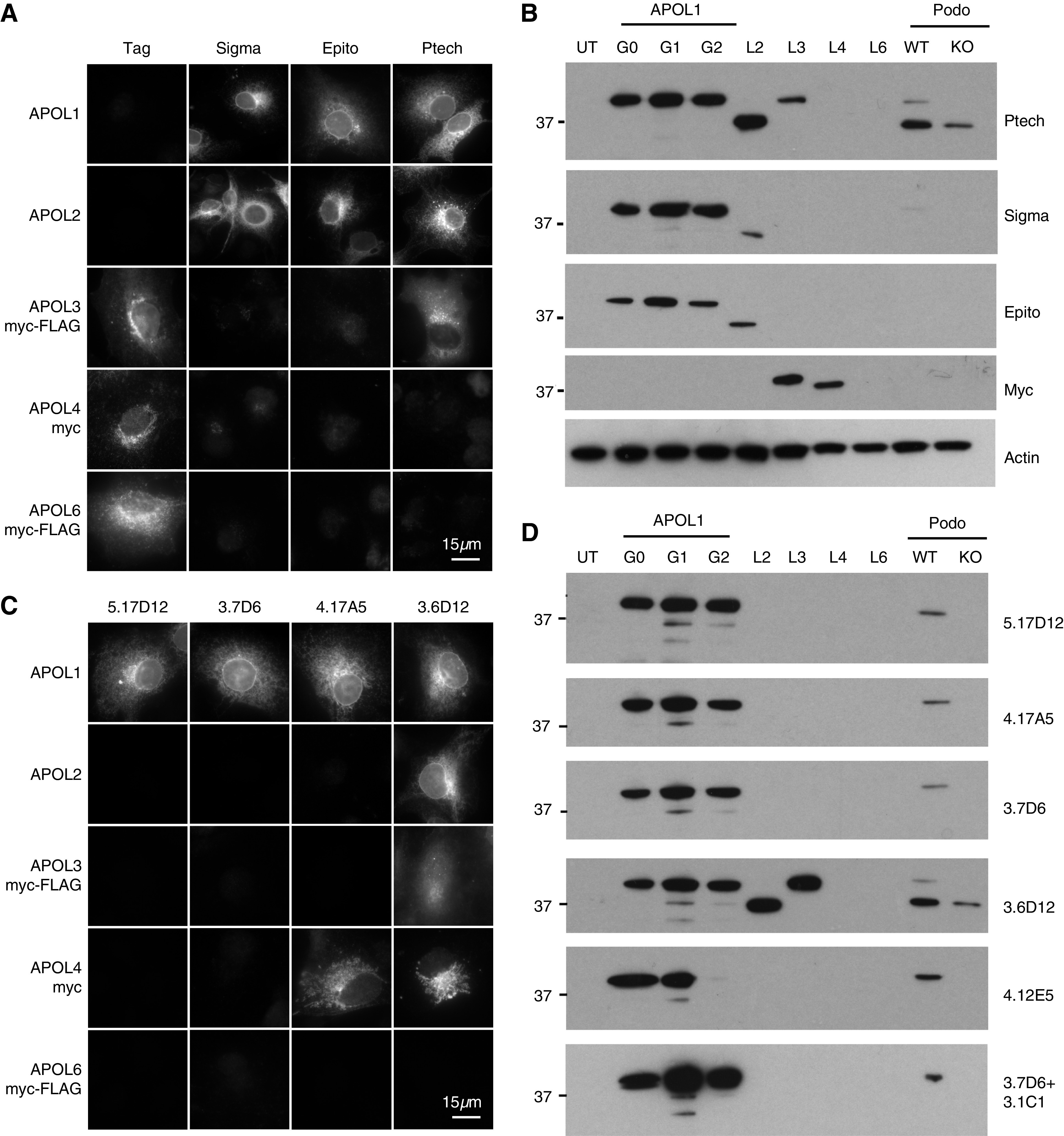
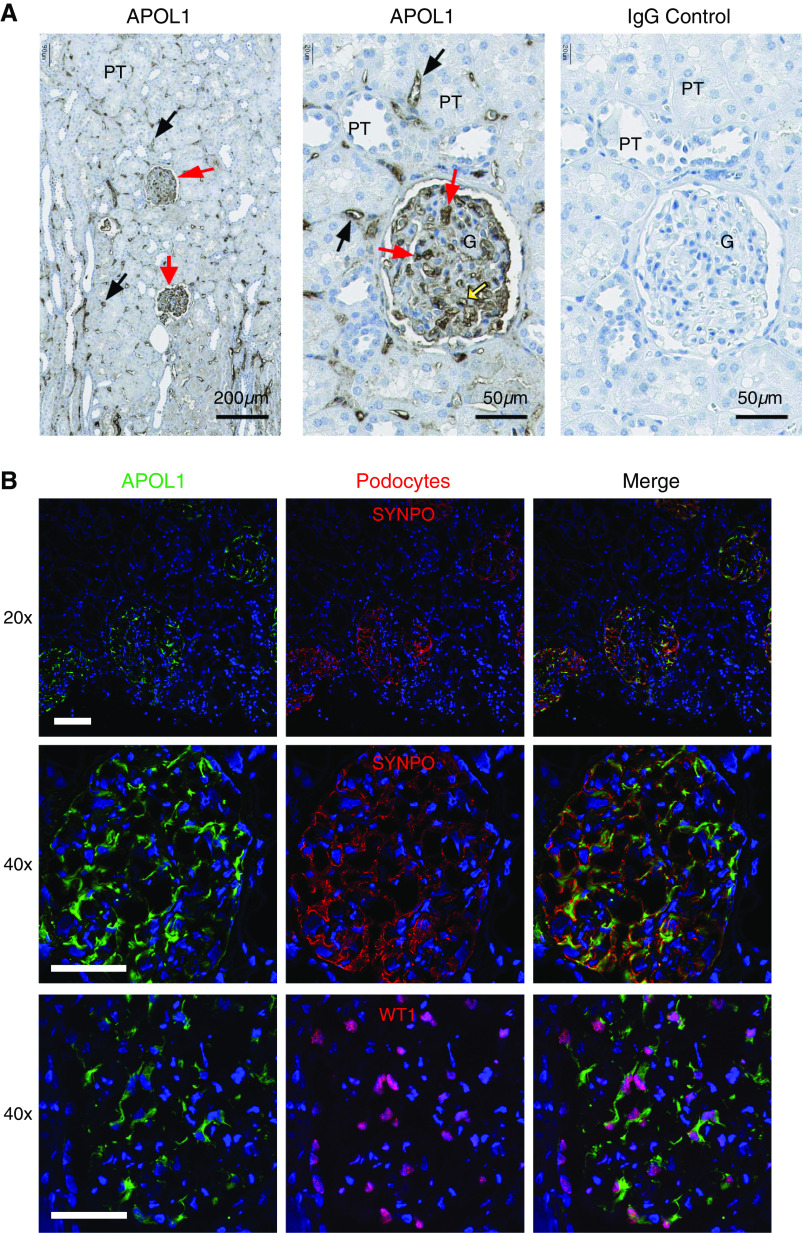
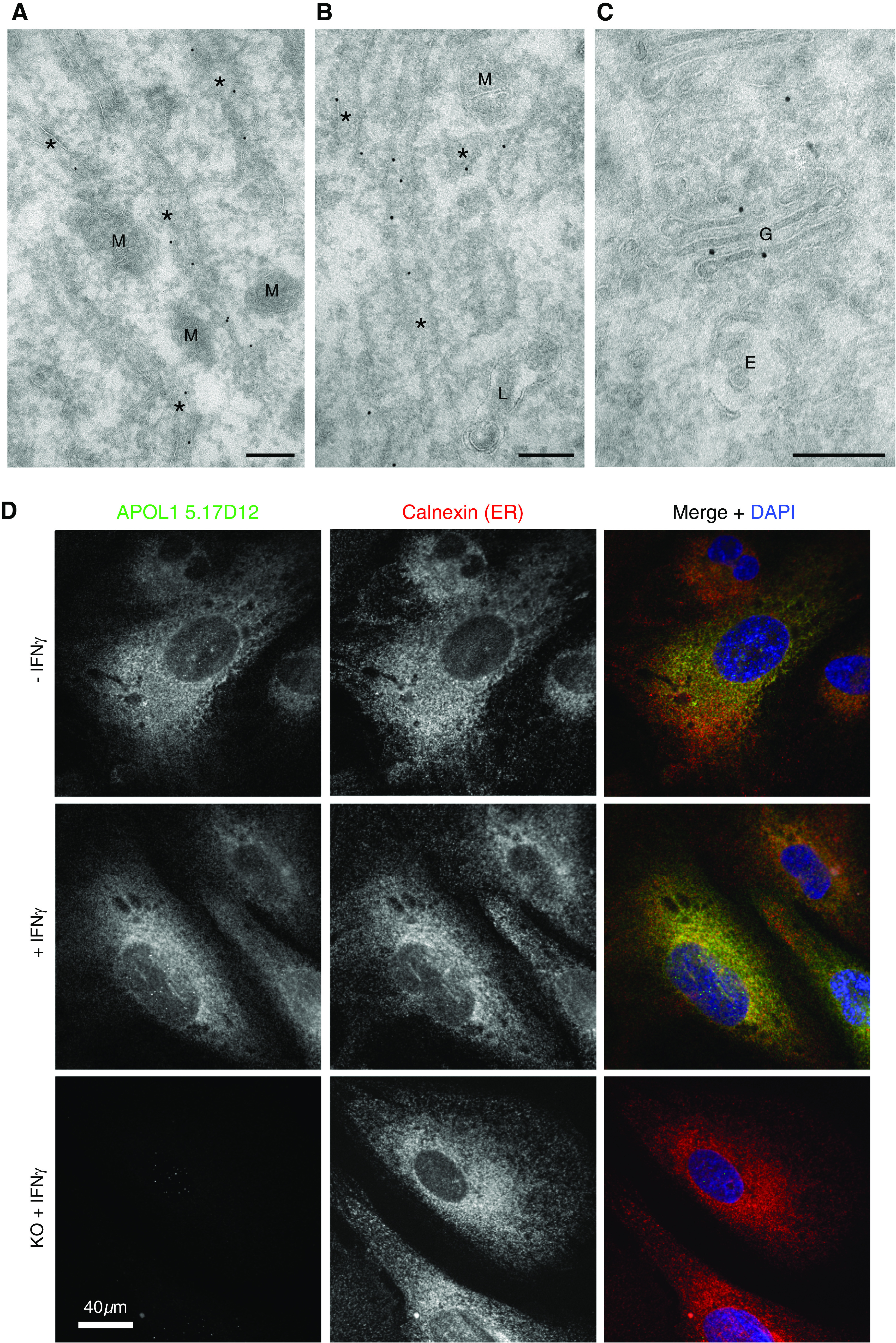
Methods: Immunohistochemistry, confocal and immunoelectron microscopy, and podocyte fractionation localized endogenous and transfected APOL1 using a large panel of novel APOL1-specific mouse and rabbit monoclonal antibodies.
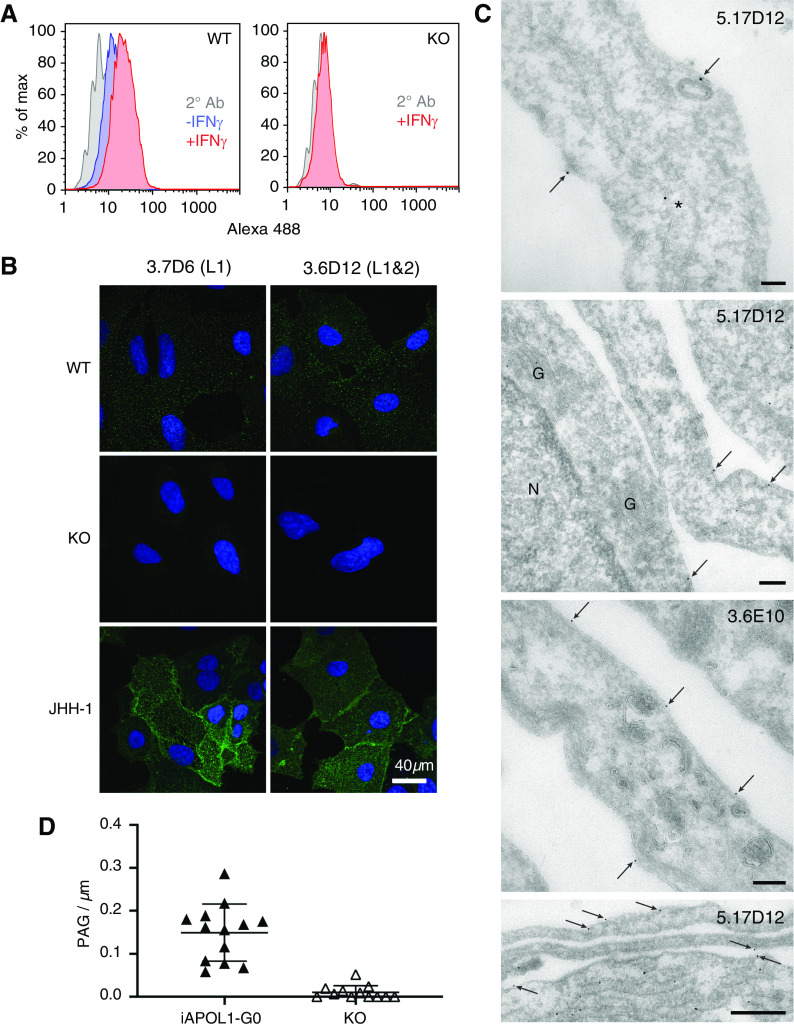
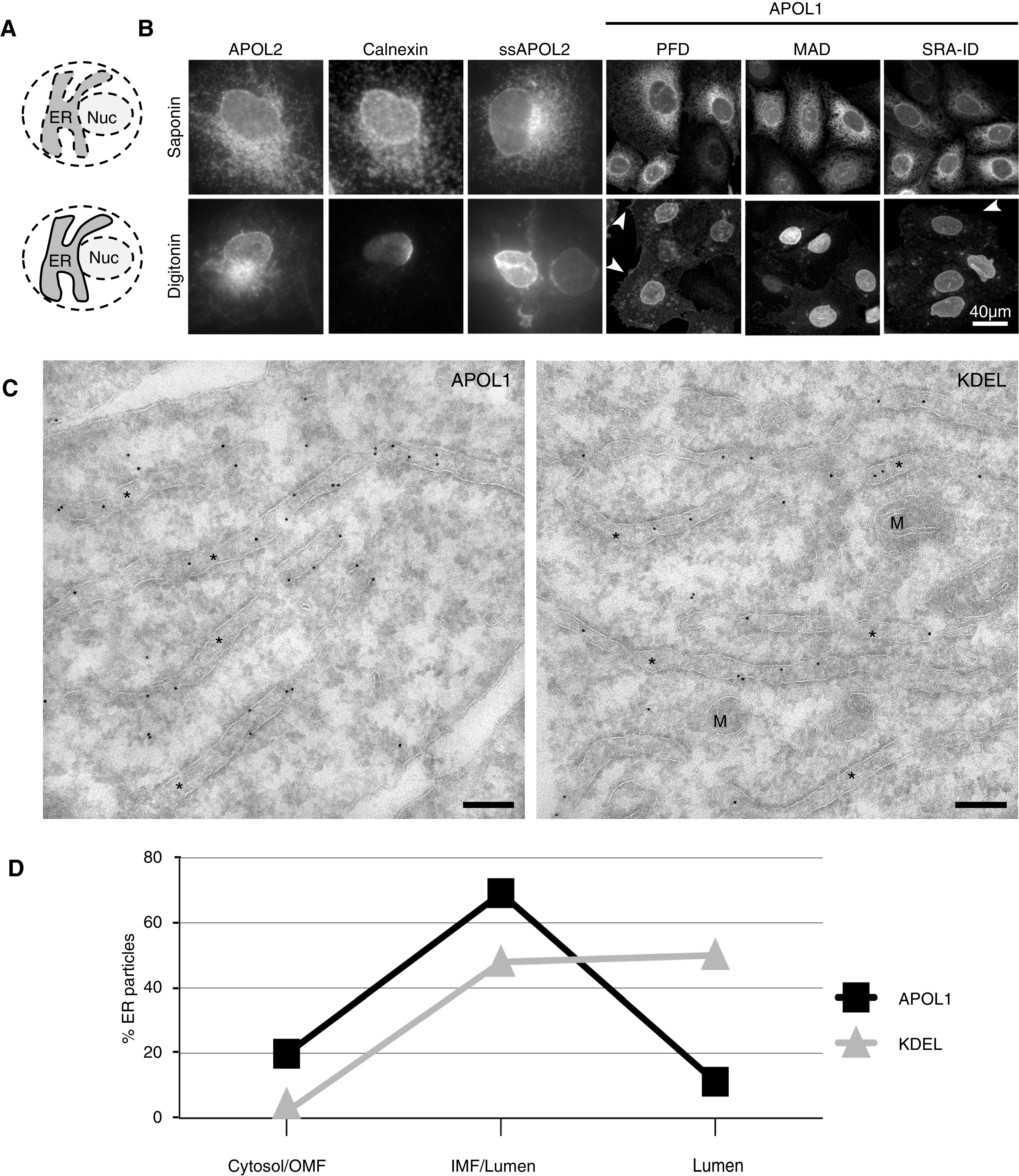
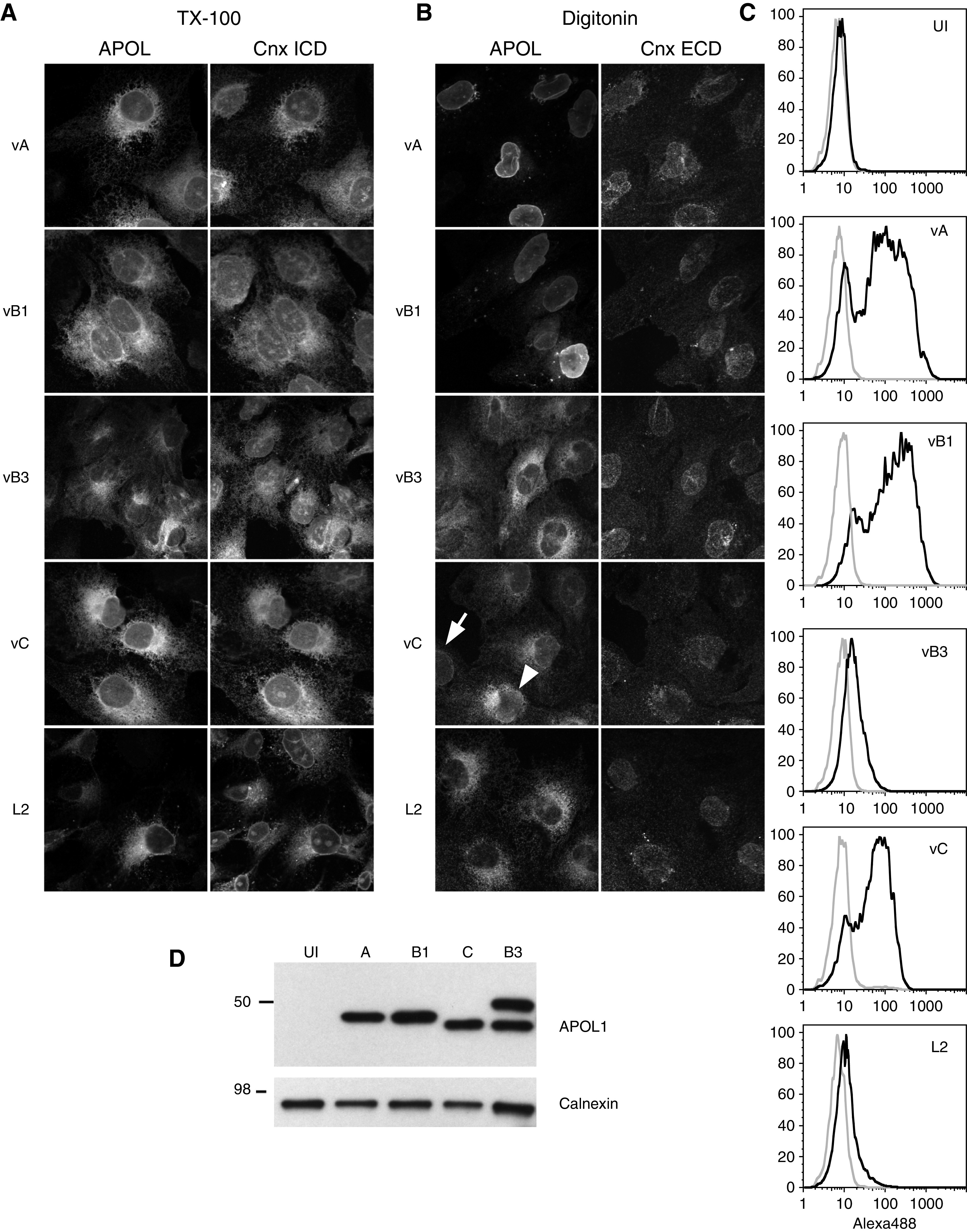
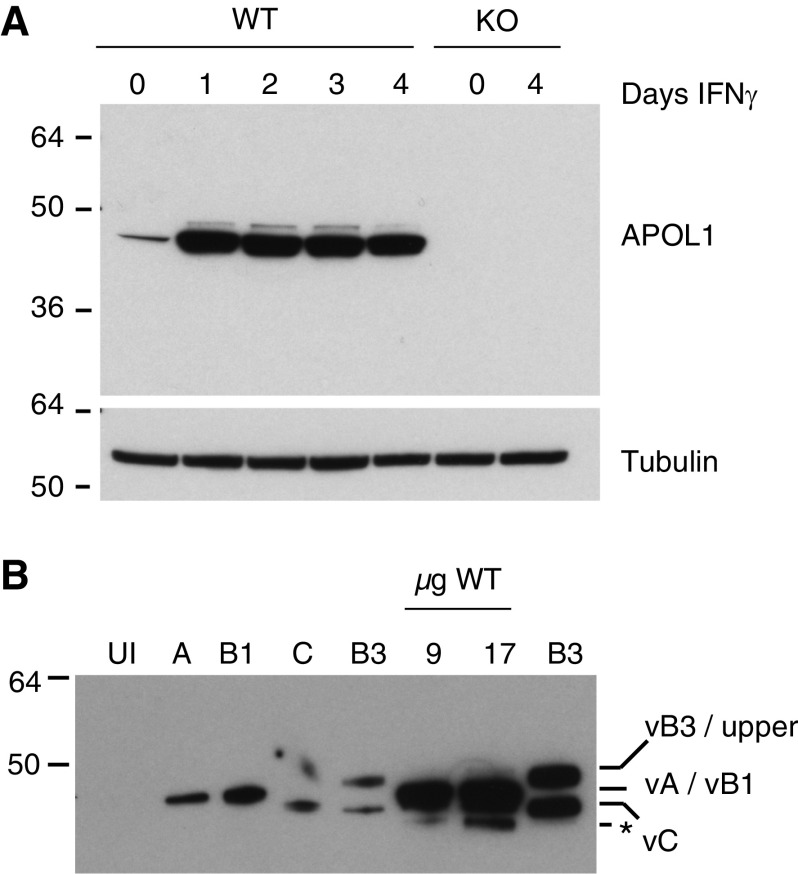
Results: Both endogenous podocyte and transfected APOL1 isoforms vA and vB1 (and a little of isoform vC) localize to the luminal face of the endoplasmic reticulum (ER) and to the cell surface, but not to mitochondria, endosomes, or lipid droplets. In contrast, APOL2, isoform vB3, and most vC of APOL1 localize to the cytoplasmic face of the ER and are consequently absent from the cell surface. APOL1 knockout podocytes do not stain for APOL1, attesting to the APOL1-specificity of the antibodies. Stable re-transfection of knockout podocytes with inducible APOL1-G0, -G1, and -G2 showed no differences in localization among variants.
Conclusions: APOL1 is found in the ER and plasma membrane, consistent with either the ER stress or surface cation channel models of APOL1-mediated cytotoxicity. The surface localization of APOL1 variants potentially opens new therapeutic targeting avenues.
Keywords: Apolipoprotein L1; Apolipoprotein L2; chronic kidney disease; immunofluorescence; immunohistochemistry; podocyte.
Copyright © 2020 by the American Society of Nephrology.
Figures
References
-
- Kruzel-Davila E, Skorecki K: Dilemmas and challenges in apolipoprotein L1 nephropathy research. Curr Opin Nephrol Hypertens 28: 77–86, 2019. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
