

Inhibition of PIKfyve kinase prevents infection by Zaire ebolavirus and SARS-CoV-2
- PMID: 32764148
- PMCID: PMC7456157
- DOI: 10.1073/pnas.2007837117
Inhibition of PIKfyve kinase prevents infection by Zaire ebolavirus and SARS-CoV-2
Abstract
Virus entry is a multistep process. It initiates when the virus attaches to the host cell and ends when the viral contents reach the cytosol. Genetically unrelated viruses can subvert analogous subcellular mechanisms and use similar trafficking pathways for successful entry. Antiviral strategies targeting early steps of infection are therefore appealing, particularly when the probability for successful interference through a common step is highest. We describe here potent inhibitory effects on content release and infection by chimeric vesicular stomatitis virus (VSV) containing the envelope proteins of Zaire ebolavirus (VSV-ZEBOV) or severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) (VSV-SARS-CoV-2) elicited by Apilimod and Vacuolin-1, small-molecule inhibitors of the main endosomal phosphatidylinositol-3-phosphate/phosphatidylinositol 5-kinase, PIKfyve. We also describe potent inhibition of SARS-CoV-2 strain 2019-nCoV/USA-WA1/2020 by Apilimod. These results define tools for studying the intracellular trafficking of pathogens elicited by inhibition of PIKfyve kinase and suggest the potential for targeting this kinase in developing small-molecule antivirals against SARS-CoV-2.
Keywords: APILIMOD; COVID-19; SARS-CoV-2; Vacuolin-1; ZEBOV.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
Competing interest statement: M.S.D. is a consultant for Inbios, Vir Biotechnology, and NGM Biopharmaceuticals, and is on the Scientific Advisory Board of Moderna. The M.S.D. laboratory at Washington University School of Medicine has received sponsored research agreements from Moderna and Emergent BioSolutions.
Figures
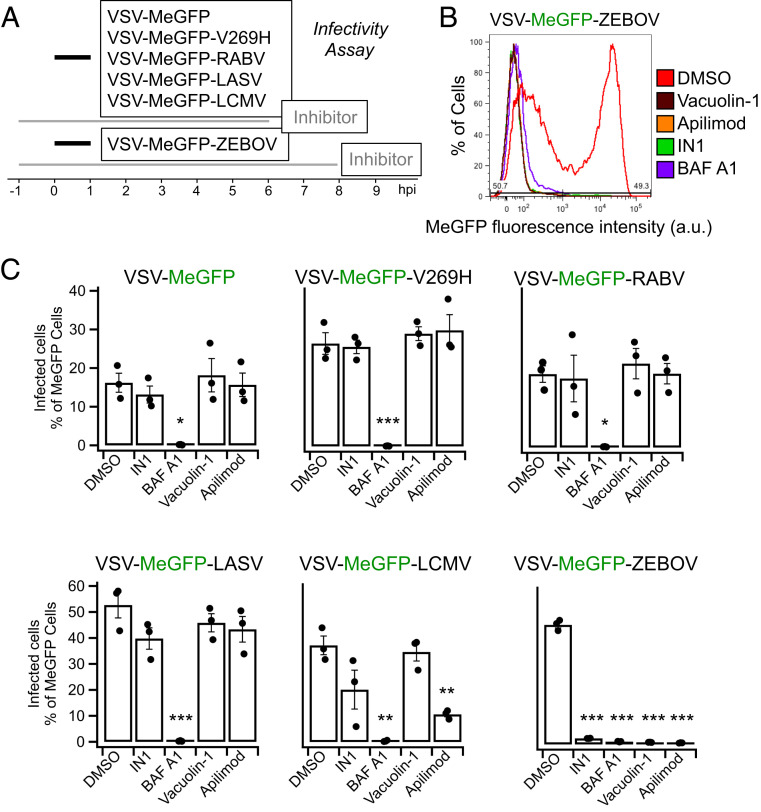
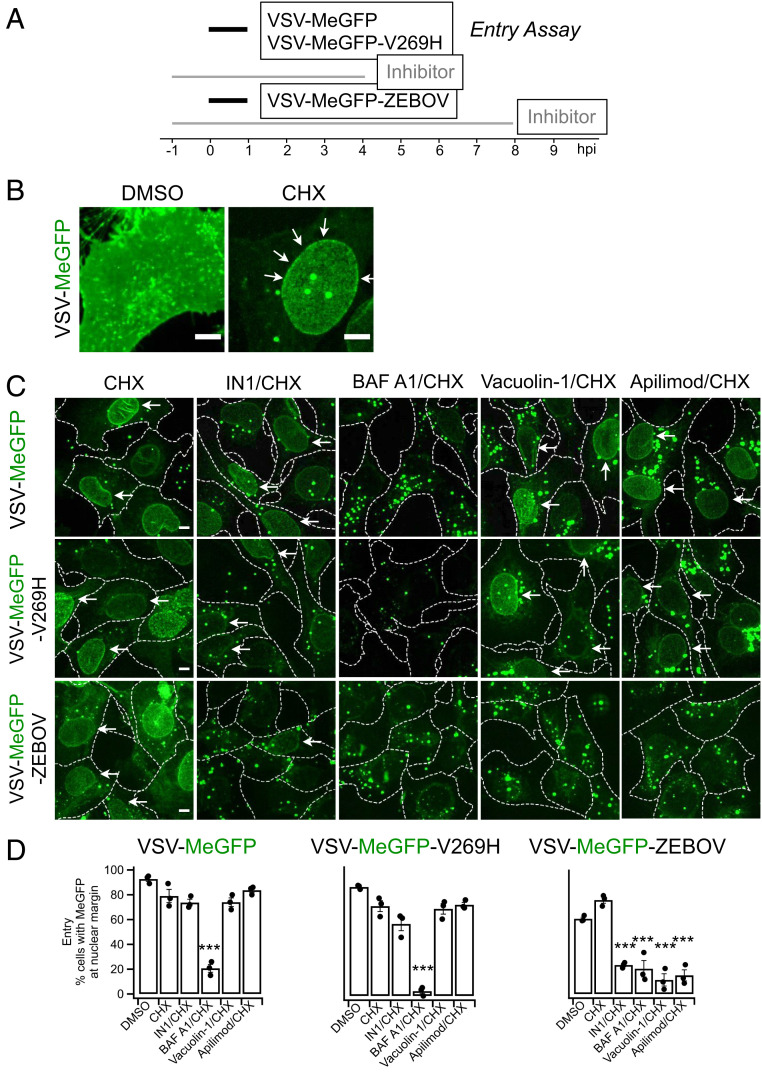
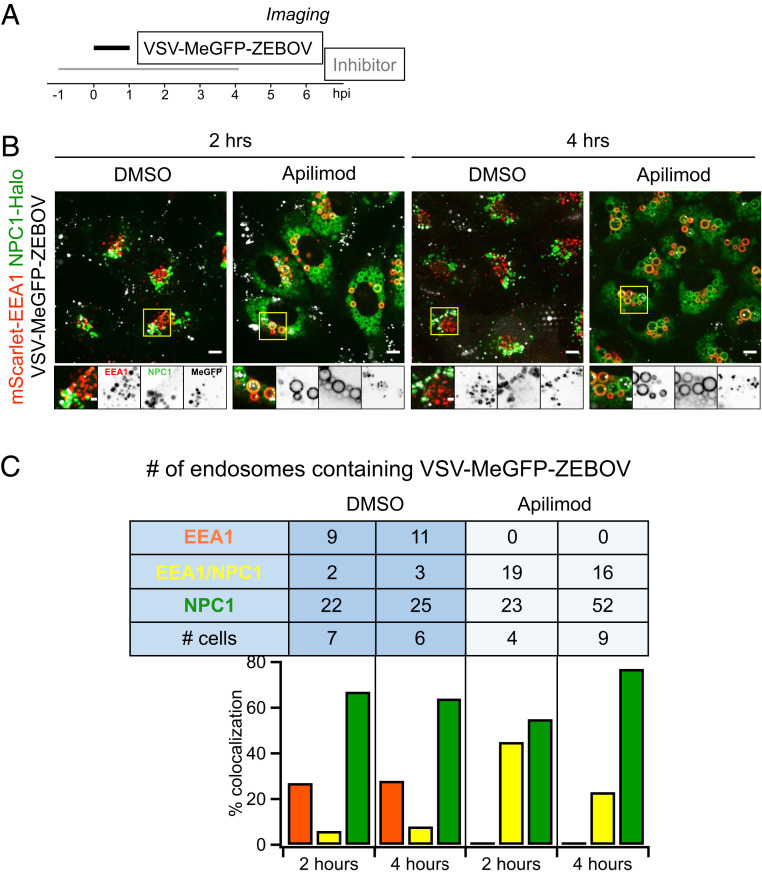
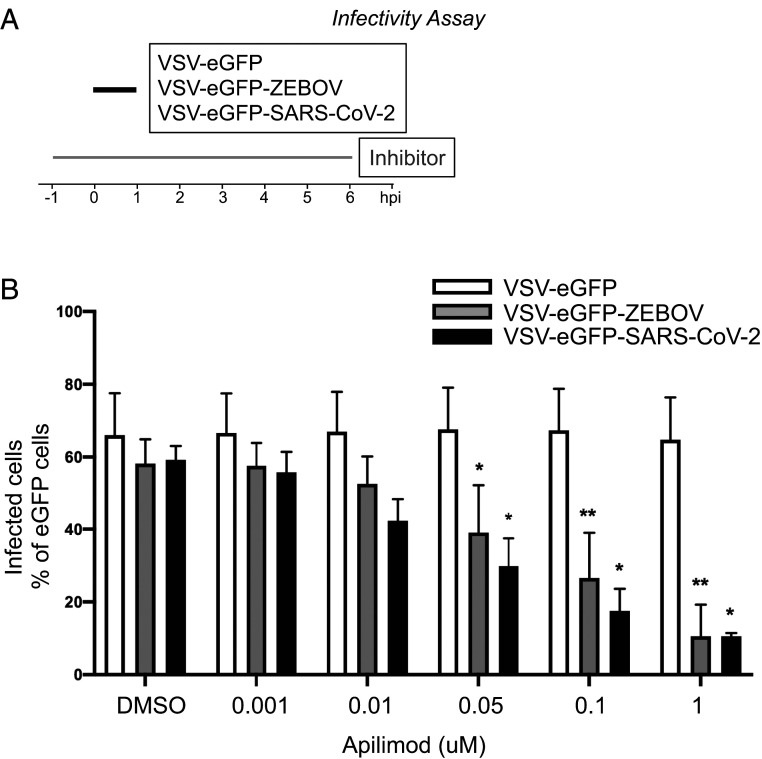
Update of
-
Inhibition of PIKfyve kinase prevents infection by Zaire ebolavirus and SARS-CoV-2.bioRxiv [Preprint]. 2020 Jun 15:2020.04.21.053058. doi: 10.1101/2020.04.21.053058. bioRxiv. 2020. Update in: Proc Natl Acad Sci U S A. 2020 Aug 25;117(34):20803-20813. doi: 10.1073/pnas.2007837117. PMID: 32511398 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
