

High leaf mass per area Oryza genotypes invest more leaf mass to cell wall and show a low mesophyll conductance
- PMID: 32765824
- PMCID: PMC7396964
- DOI: 10.1093/aobpla/plaa028
High leaf mass per area Oryza genotypes invest more leaf mass to cell wall and show a low mesophyll conductance
Abstract
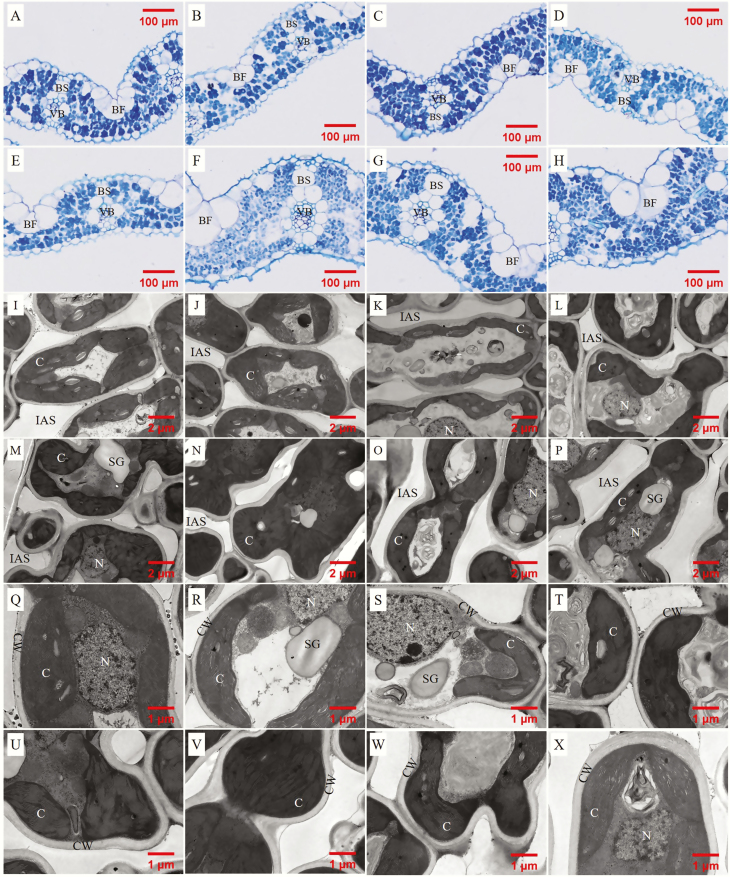
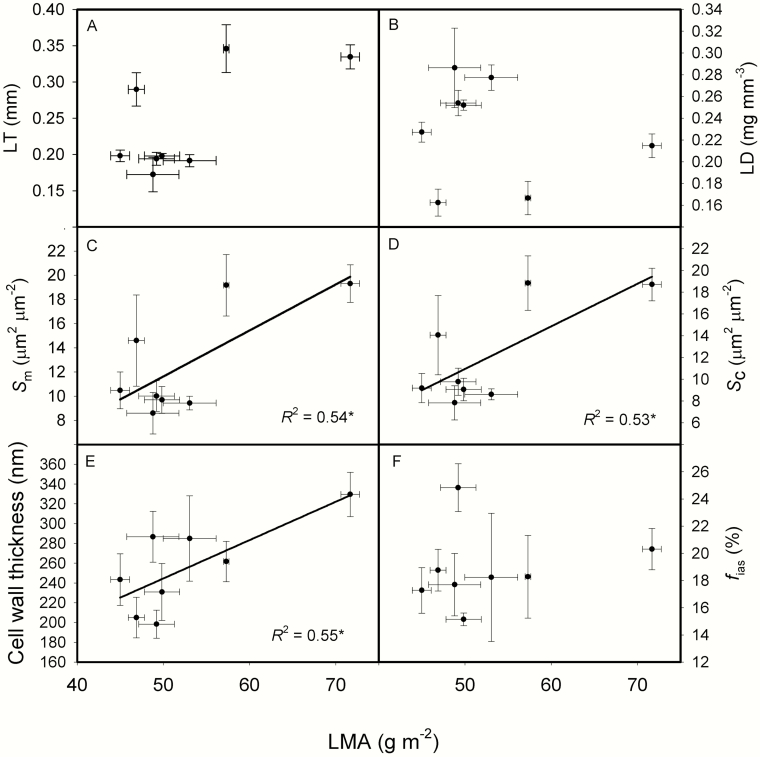
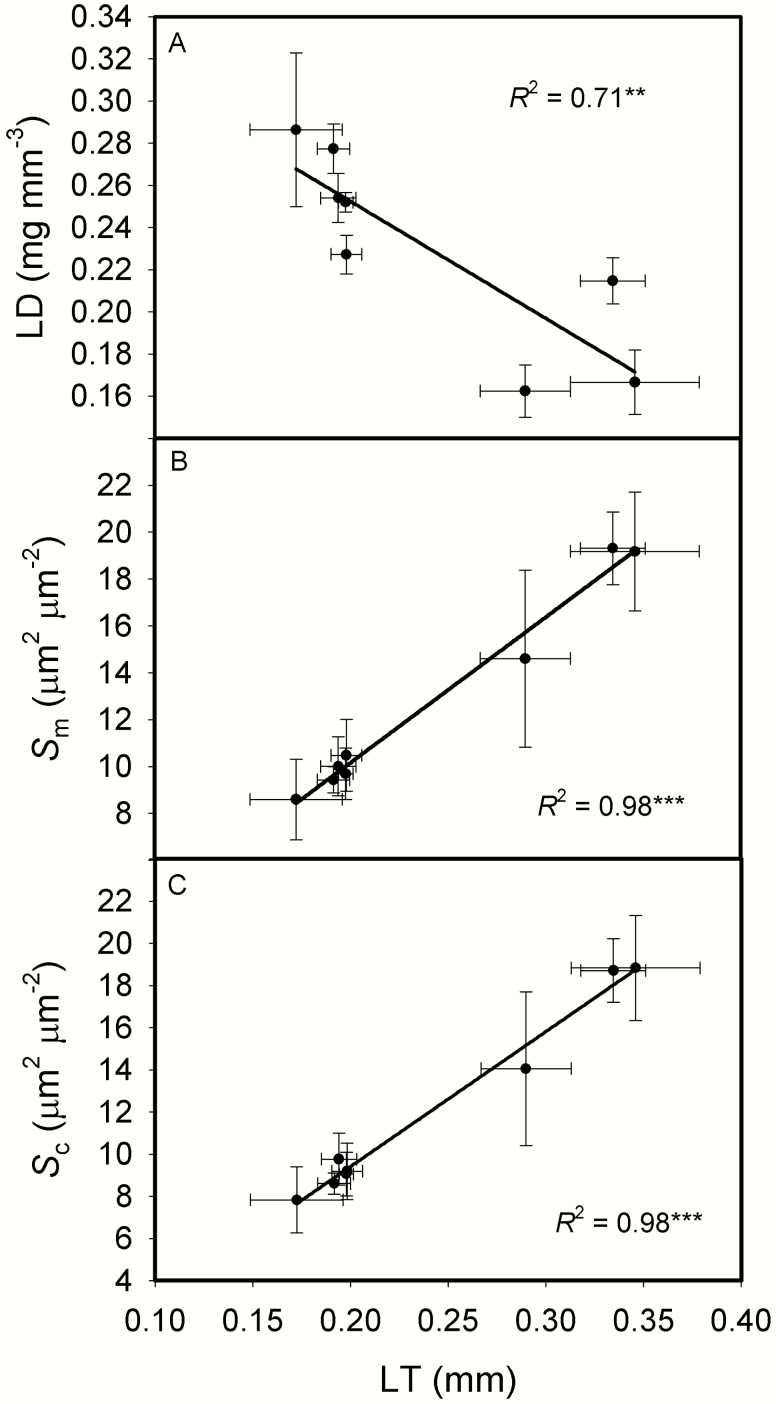
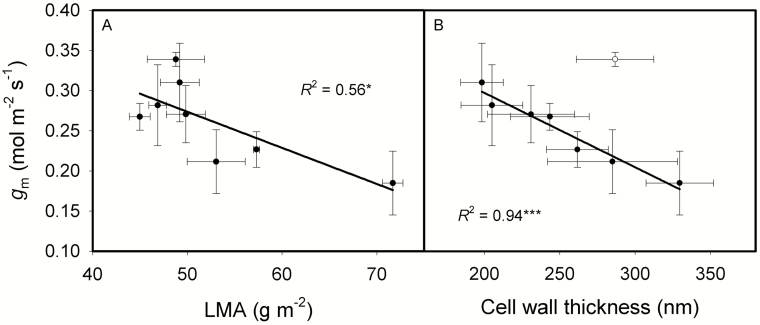
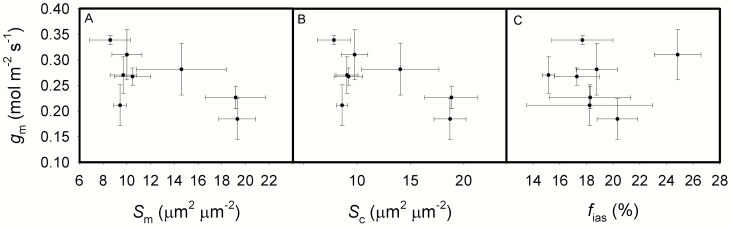
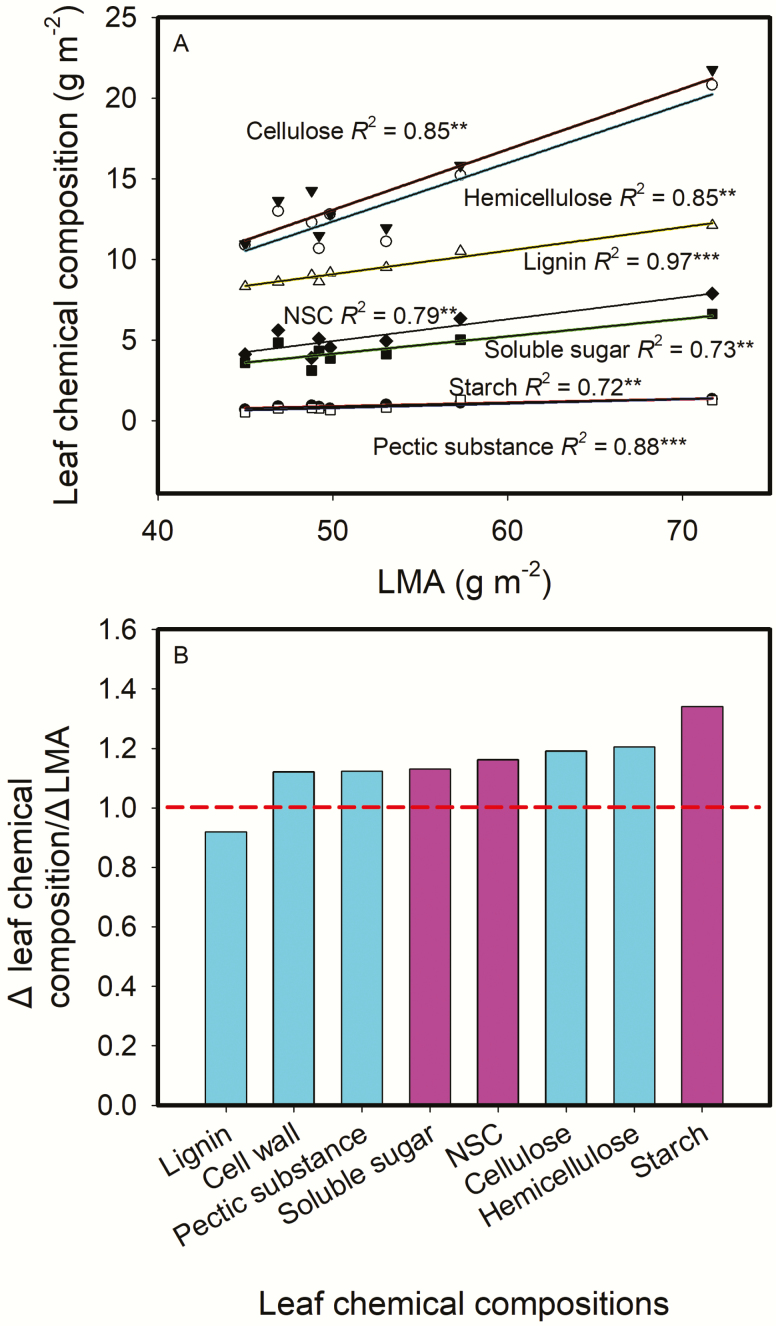
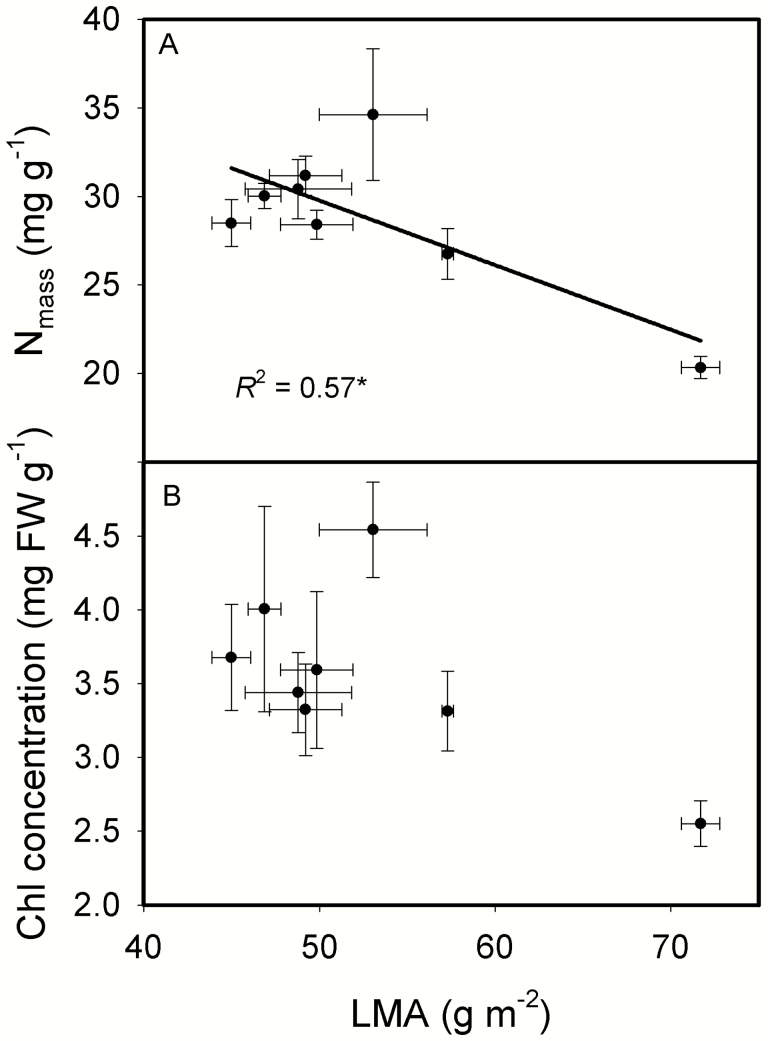
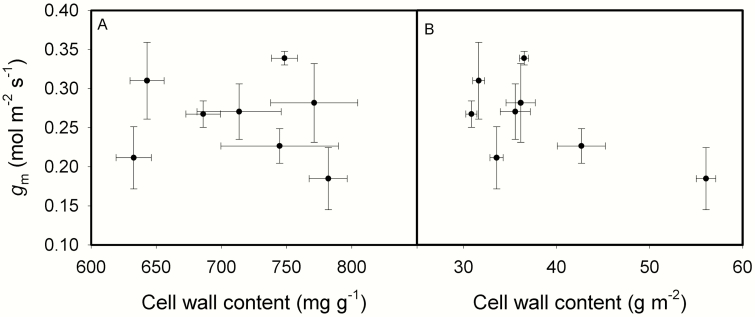
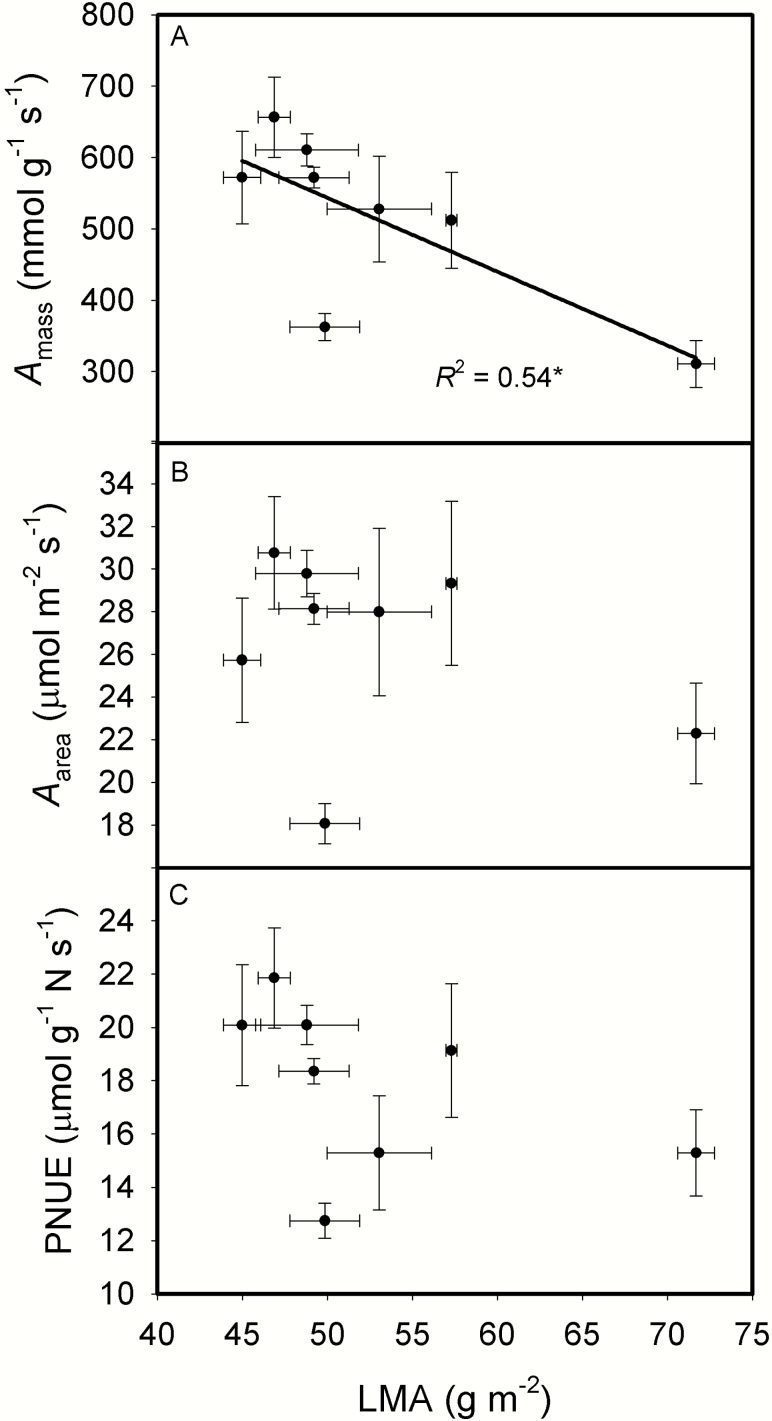
The intraspecific variations of leaf structure and anatomy in rice leaves and their impacts on gas diffusion are still unknown. Researches about the tradeoff between structural compositions and intracellular chemical components within rice leaves are still lacking. The objectives of the present study were to investigate the varietal differences in leaf structure and leaf chemical compositions, and the tradeoff between leaf structural tissues and intracellular chemical components in rice leaves. Leaf structure, leaf anatomy, leaf chemical composition concentrations and gas exchange parameters were measured on eight Oryza sativa L. genotypes to investigate the intraspecific variations in leaf structure and leaf anatomy and their impacts on gas exchange parameters, and to study the tradeoff between leaf structural compositions (cell wall compounds) and intracellular chemical components (non-structural carbohydrates, nitrogen, chlorophyll). Leaf thickness increased with leaf mass per area (LMA), while leaf density did not correlate with LMA. Mesophyll cell surface area exposed to intercellular airspace (IAS) per leaf area, the surface area of chloroplasts exposed to IAS and cell wall thickness increased with LMA. Cell wall compounds accounted for 71.5 % of leaf dry mass, while mass-based nitrogen and chlorophyll concentrations decreased with LMA. Mesophyll conductance was negatively correlated with LMA and cell wall thickness. High LMA rice genotypes invest more leaf mass to cell wall and possess a low mesophyll conductance.
Keywords: Cell wall; Oryza sativa L; leaf anatomy; leaf chemical compositions; leaf mass per area; tradeoff.
© The Author(s) 2020. Published by Oxford University Press on behalf of the Annals of Botany Company.
Figures
References
-
- Adachi S, Nakae T, Uchida M, Soda K, Takai T, Oi T, Yamamoto T, Ookawa T, Miyake H, Yano M, Hirasawa T. 2013. The mesophyll anatomy enhancing CO2 diffusion is a key trait for improving rice photosynthesis. Journal of Experimental Botany 64:1061–1072. - PubMed
-
- Brooks A, Farquhar GD. 1985. Effect of temperature on the CO2/O2 specificity of ribulose-1,5-bisphosphate carboxylase/oxygenase and the rate of respiration in the light: estimates from gas-exchange measurements on spinach. Planta 165:397–406. - PubMed
-
- Castro-Díez P, Puyravaud JP, Cornelissen JH. 2000. Leaf structure and anatomy as related to leaf mass per area variation in seedlings of a wide range of woody plant species and types. Oecologia 124:476–486. - PubMed
-
- Choong MF, Lucas PW, Ong JSY, Pereira B, Tan HTW, Turner IM. 1992. Leaf fracture toughness and sclerophylly: their correlations and ecological implications. New Phytologist 121:597–610.
-
- Coley PD. 1983. Herbivory and defensive characteristics of tree species in a low land tropical forest. Ecological Monographs 53:209–233.
LinkOut - more resources
Full Text Sources
