

Chemotaxis strategies of bacteria with multiple run modes
- PMID: 32766440
- PMCID: PMC7385427
- DOI: 10.1126/sciadv.aaz6153
Chemotaxis strategies of bacteria with multiple run modes
Abstract
Bacterial chemotaxis-a fundamental example of directional navigation in the living world-is key to many biological processes, including the spreading of bacterial infections. Many bacterial species were recently reported to exhibit several distinct swimming modes-the flagella may, for example, push the cell body or wrap around it. How do the different run modes shape the chemotaxis strategy of a multimode swimmer? Here, we investigate chemotactic motion of the soil bacterium Pseudomonas putida as a model organism. By simultaneously tracking the position of the cell body and the configuration of its flagella, we demonstrate that individual run modes show different chemotactic responses in nutrition gradients and, thus, constitute distinct behavioral states. On the basis of an active particle model, we demonstrate that switching between multiple run states that differ in their speed and responsiveness provides the basis for robust and efficient chemotaxis in complex natural habitats.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures

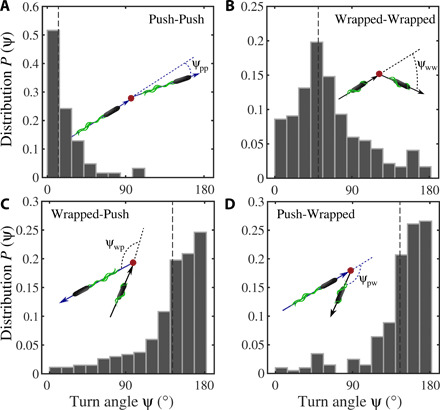
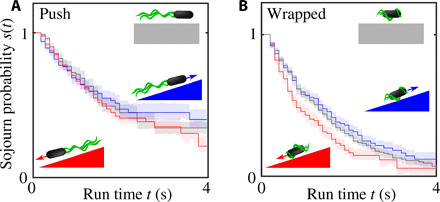
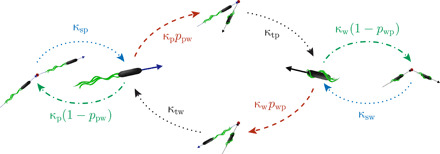
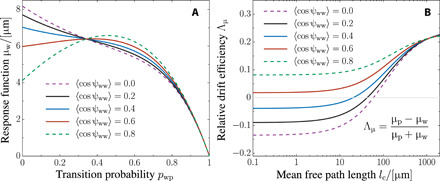
References
-
- H. C. Berg, E. coli in Motion (Springer, 2004).
-
- Berg H. C., Brown D. A., Chemotaxis in Escherichia coli analysed by three-dimensional tracking. Nature 239, 500–504 (1972). - PubMed
-
- Johansen J. E., Pinhassi J., Blackburn N., Zweifel U. L., Hagström Å., Variability in motility characteristics among marine bacteria. Aquat. Microb. Ecol. 28, 229–237 (2002).
Publication types
LinkOut - more resources
Full Text Sources
