

The inhibition of cellular toxicity of amyloid-β by dissociated transthyretin
- PMID: 32769117
- PMCID: PMC7549042
- DOI: 10.1074/jbc.RA120.013440
The inhibition of cellular toxicity of amyloid-β by dissociated transthyretin
Abstract
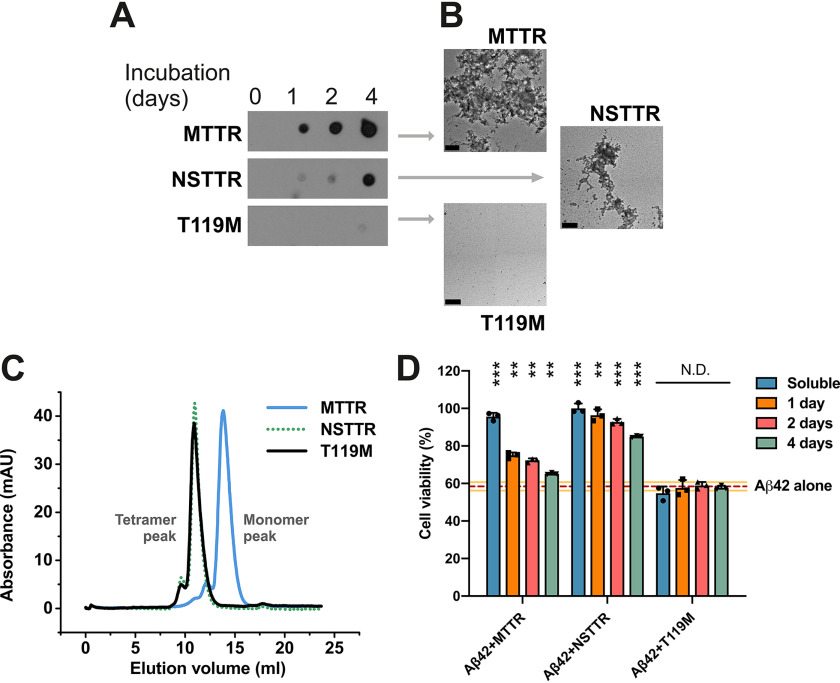
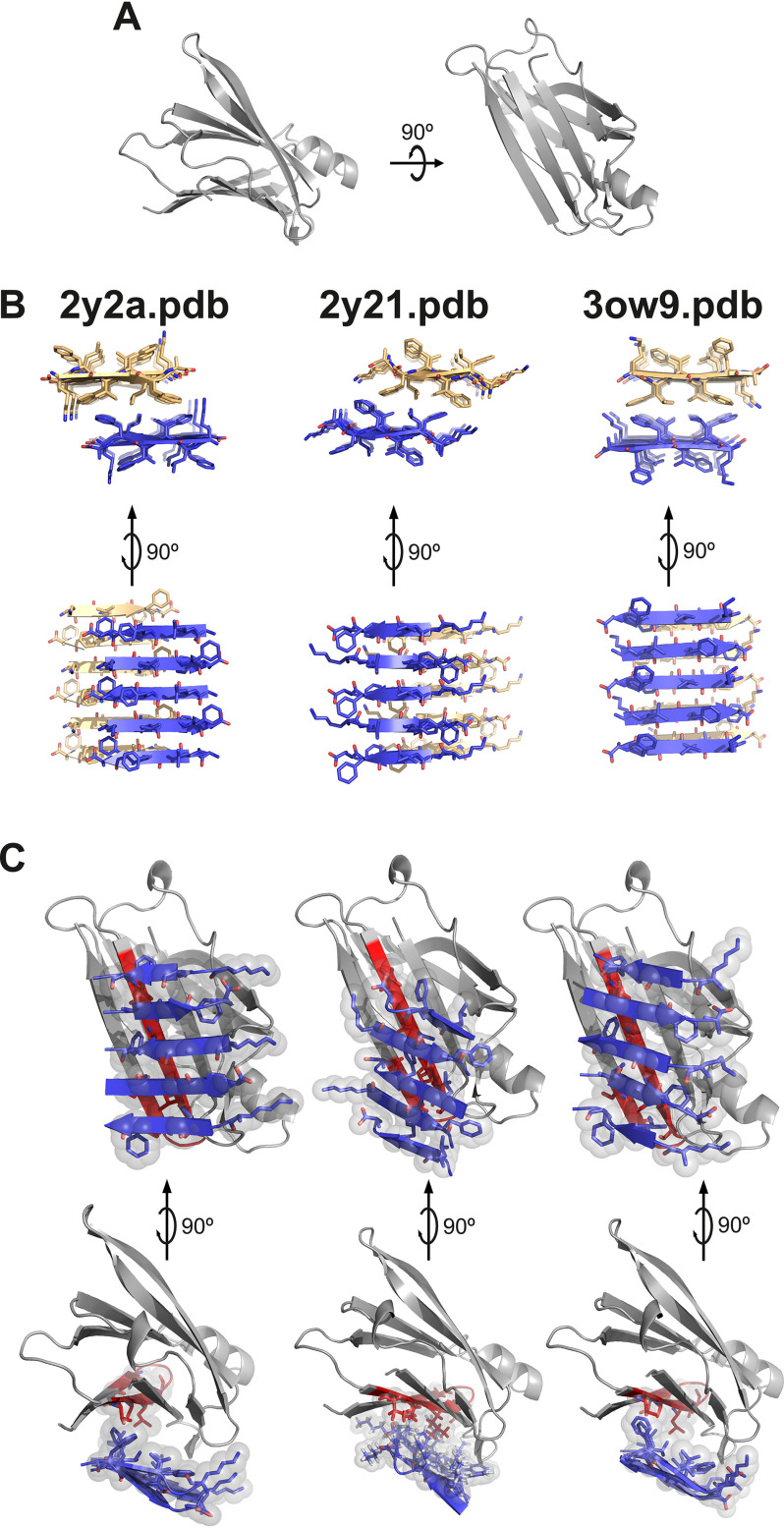
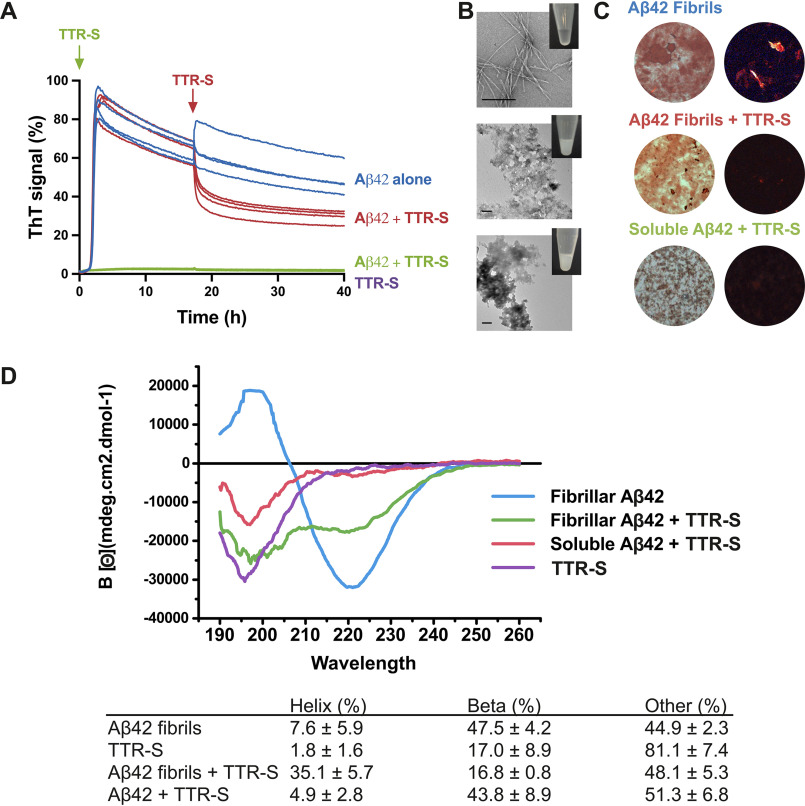
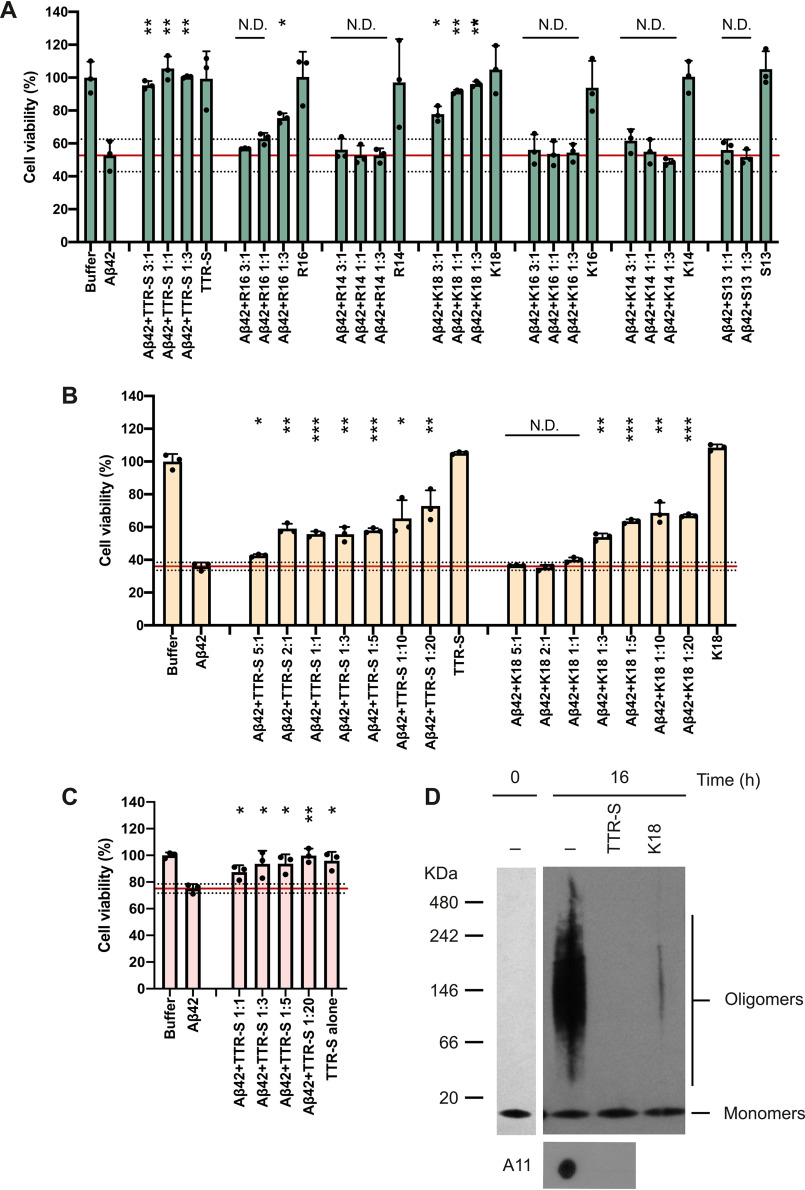
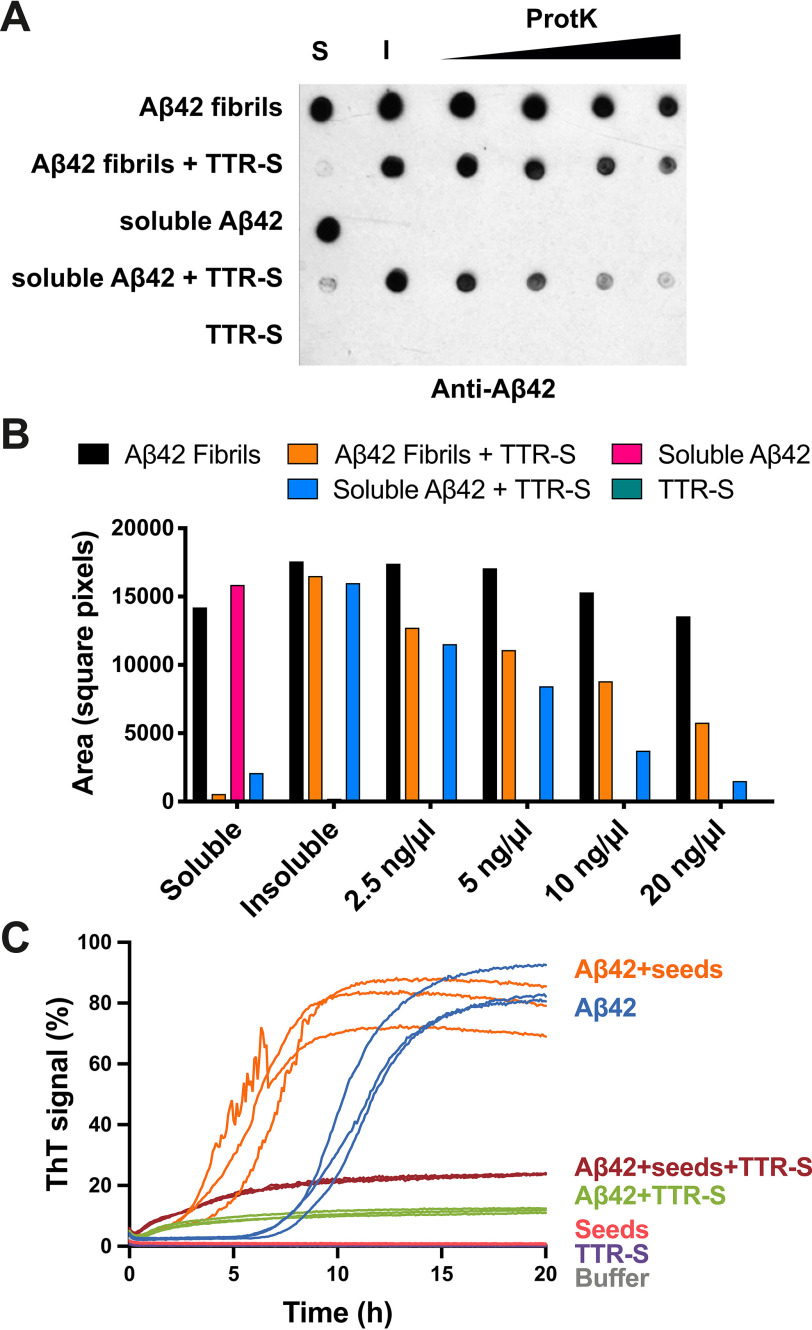
The protective effect of transthyretin (TTR) on cellular toxicity of β-amyloid (Aβ) has been previously reported. TTR is a tetrameric carrier of thyroxine in blood and cerebrospinal fluid, the pathogenic aggregation of which causes systemic amyloidosis. However, studies have documented a protective effect of TTR against cellular toxicity of pathogenic Aβ, a protein associated with Alzheimer's disease. TTR binds Aβ, alters its aggregation, and inhibits its toxicity both in vitro and in vivo In this study, we investigate whether the amyloidogenic ability of TTR and its antiamyloid inhibitory effect are associated. Using protein aggregation and cytotoxicity assays, we found that the dissociation of the TTR tetramer, required for its amyloid pathogenesis, is also necessary to prevent cellular toxicity from Aβ oligomers. These findings suggest that the Aβ-binding site of TTR may be hidden in its tetrameric form. Aided by computational docking and peptide screening, we identified a TTR segment that is capable of altering Aβ aggregation and toxicity, mimicking TTR cellular protection. EM, immune detection analysis, and assessment of aggregation and cytotoxicity revealed that the TTR segment inhibits Aβ oligomer formation and also promotes the formation of nontoxic, nonamyloid amorphous aggregates, which are more sensitive to protease digestion. Finally, this segment also inhibits seeding of Aβ catalyzed by Aβ fibrils extracted from the brain of an Alzheimer's patient. Together, these findings suggest that mimicking the inhibitory effect of TTR with peptide-based therapeutics represents an additional avenue to explore for the treatment of Alzheimer's disease.
Keywords: Alzheimer's disease; amyloid; amyloid-beta (AB); cytotoxicity; neuroprotection; neuroscience; peptide; transthyretin.
© 2020 Cao et al.
Conflict of interest statement
Conflict of interest—L. S. is a consultant for ADRx, Inc.
Figures
References
-
- Schwarzman A. L., Gregori L., Vitek M. P., Lyubski S., Strittmatter W. J., Enghilde J. J., Bhasin R., Silverman J., Weisgraber K. H., Coyle P. K. (1994) Transthyretin sequesters amyloid β protein and prevents amyloid formation. Proc. Natl. Acad. Sci. U.S.A. 91, 8368–8372 10.1073/pnas.91.18.8368 - DOI - PMC - PubMed
-
- Roher A. E., Lowenson J. D., Clarke S., Woods A. S., Cotter R. J., Gowing E., and Ball M. J. (1993) β-Amyloid-(1–42) is a major component of cerebrovascular amyloid deposits: implications for the pathology of Alzheimer disease. Proc. Natl. Acad. Sci. U.S.A. 90, 10836–10840 10.1073/pnas.90.22.10836 - DOI - PMC - PubMed
-
- Burdick D., Soreghan B., Kwon M., Kosmoski J., Knauer M., Henschen A., Yates J., Cotman C., and Glabe C. (1992) Assembly and aggregation properties of synthetic Alzheimer's A4/β amyloid peptide analogs. J. Biol. Chem. 267, 546–554 - PubMed
-
- Stein T. D., Anders N. J., DeCarli C., Chan S. L., Mattson M. P., and Johnson J. A. (2004) Neutralization of transthyretin reverses the neuroprotective effects of secreted amyloid precursor protein (APP) in APPSW mice resulting in tau phosphorylation and loss of hippocampal neurons: support for the amyloid hypothesis. J. Neurosci. 24, 7707–7717 10.1523/JNEUROSCI.2211-04.2004 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
