

Triacylglycerols sequester monotopic membrane proteins to lipid droplets
- PMID: 32769983
- PMCID: PMC7414839
- DOI: 10.1038/s41467-020-17585-8
Triacylglycerols sequester monotopic membrane proteins to lipid droplets
Abstract
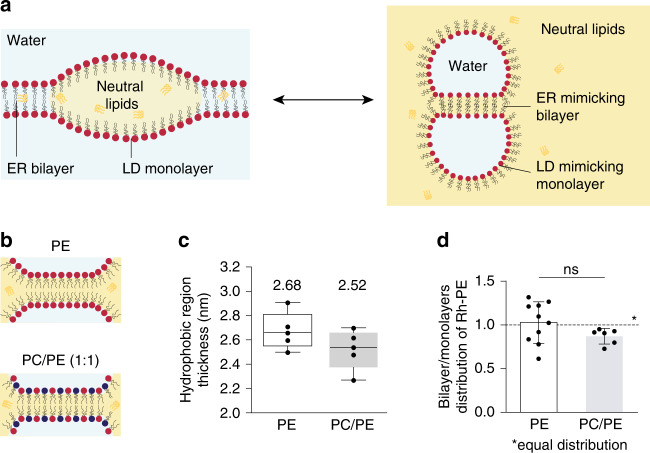
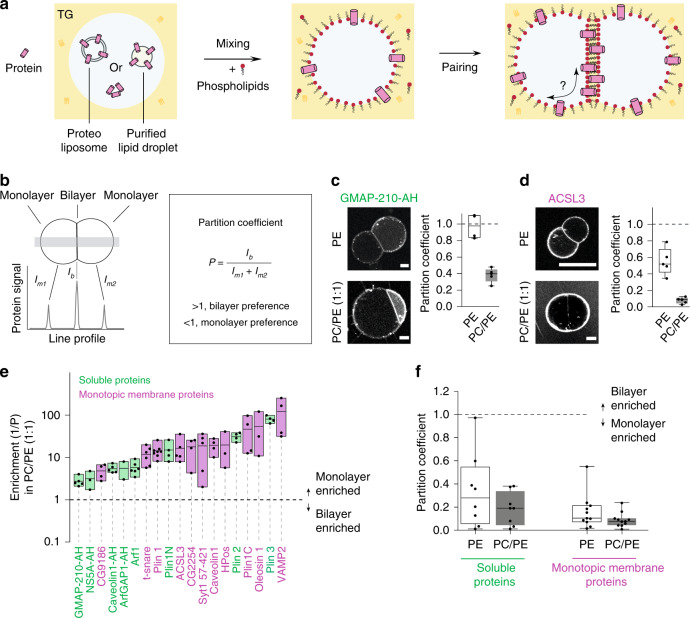
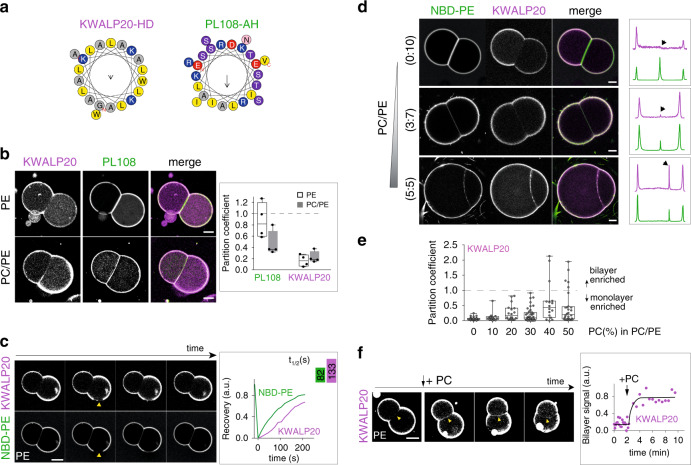
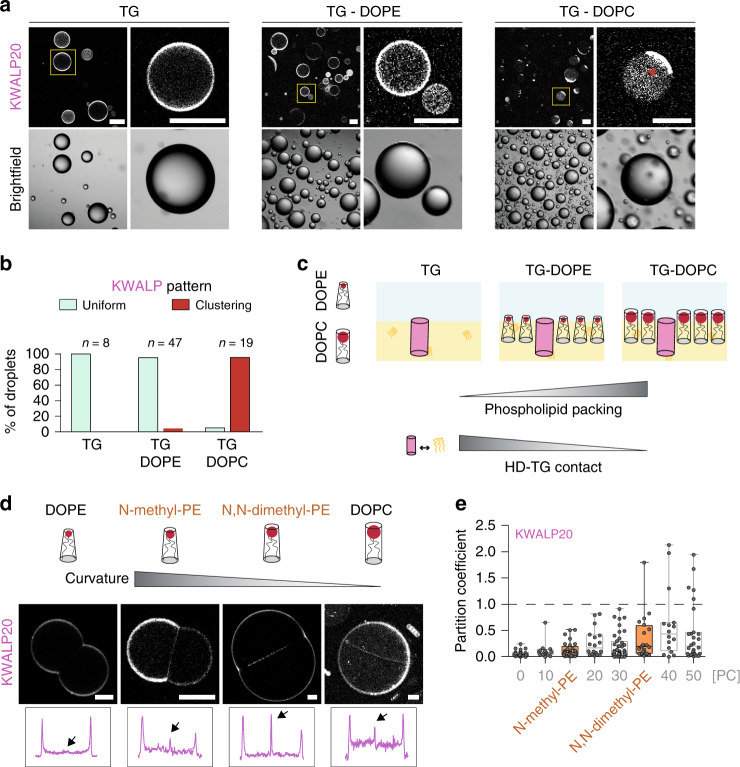
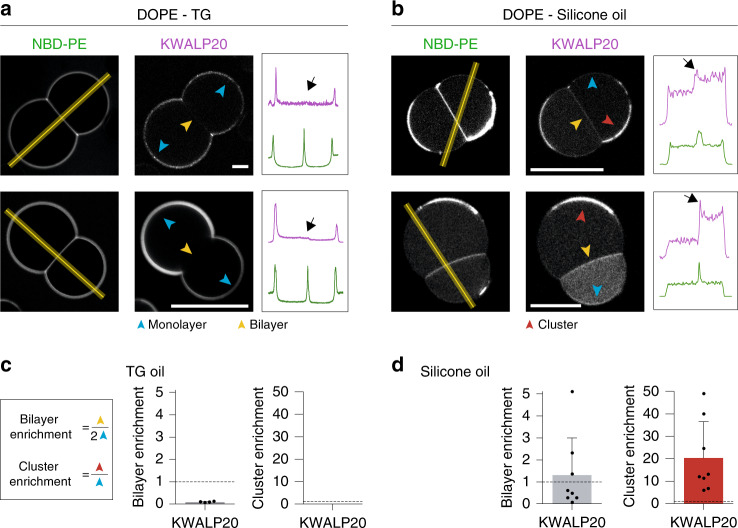
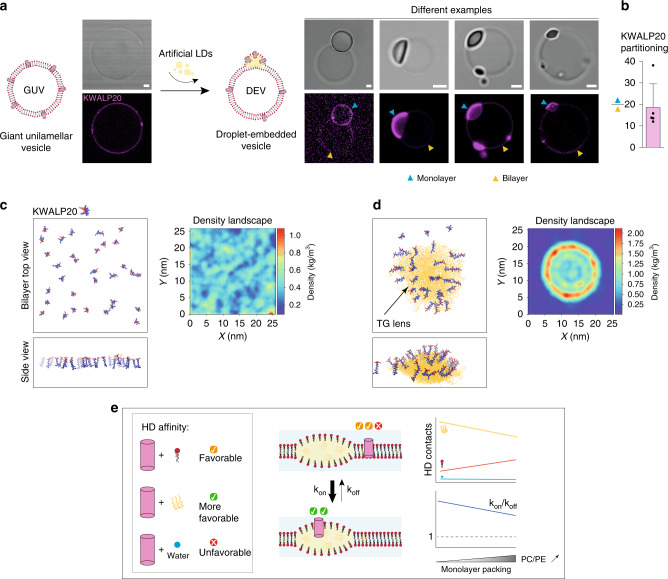
Triacylglycerols (TG) are synthesized at the endoplasmic reticulum (ER) bilayer and packaged into organelles called lipid droplets (LDs). LDs are covered by a single phospholipid monolayer contiguous with the ER bilayer. This connection is used by several monotopic integral membrane proteins, with hydrophobic membrane association domains (HDs), to diffuse between the organelles. However, how proteins partition between ER and LDs is not understood. Here, we employed synthetic model systems and found that HD-containing proteins strongly prefer monolayers and returning to the bilayer is unfavorable. This preference for monolayers is due to a higher affinity of HDs for TG over membrane phospholipids. Protein distribution is regulated by PC/PE ratio via alterations in monolayer packing and HD-TG interaction. Thus, HD-containing proteins appear to non-specifically accumulate to the LD surface. In cells, protein editing mechanisms at the ER membrane would be necessary to prevent unspecific relocation of HD-containing proteins to LDs.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Santinho A, et al. Membrane Curvature Catalyzes Lipid Droplet Assembly. Curr. Biol. 2020;30:2481–2494.e6. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
